Der Eucyt – die Zelle der Eukaryoten
Der zelluläre Baustein aller Eukaryoten, gleichgültig ob Ein- oder Vielzeller, Pflanzen, Pilze oder Tiere, ist der Eucyt. Gegenüber dem Procyten zeichnet sich dieser Zelltyp grundsätzlich in erster Linie durch folgende Baueigenheiten aus:
| |
|
|
|
|
|
|
Das Cytoplasma ist die „Grundsubstanz“ der lebenden Zelle. Es umgibt die Zellorganellen und wird auch definiert als der Bereich zwischen dem Zellkern und der die Zelle umgebenden Membran.
Das Cytoplasma besteht zu einem beachtlichen Teil aus Wasser, enthält Proteine, lösliche Kohlenhydrate und Nucleinsäuren. Auch organische Säuren und Ionen sind im Cytoplasma zu finden. Die chemische Zusammensetzung des Cytoplasmas hängt sehr vom physiologischen Zustand der Zelle, der Spezialisierung und ihrem Alter ab. Hinsichtlich der Konsistenz kann das halbflüssige Cytosol, in dem die Organellen liegen, vom dünnflüssigen Sol- in den eher zähflüssigen Gelzustand wechseln. Auffällige Bewegungen des Plasmas und der von ihm eingeschlossenen Organellen sind die Rotation, die Zirkulation und die Pulsation.
Ursprünglich nahm man an, das Cytoplasma sei eine homogene, strukturlose Flüssigkeit. Mittels Immunfluoreszenz und Elektronenmikroskopie fand man aber heraus, dass es eine ausgeprägte innere Architektur besitzt. Die aus Proteinen bestehenden Filamente und Mikrotubuli bilden ein Gerüst, das man Cytoskelett nennt.
Über die Funktionen des Cytoskeletts besteht noch keine vollkommene Klarheit. Neben der Stabilisierung der Zellgestalt dürfte es für Bewegungs-, Transport- und Informationsvorgänge mitverantwortlich sein und auch eine Rolle bei der Zellteilung spielen.
Der Zellkern (Nucleus, Karyon) gehört zu den größten Zellorganellen. Er ist meist von kugelförmiger Gestalt, in der Zelle in Einzahl vorhanden und bereits im Lichtmikroskop gut zu sehen. Der Kern wird von einer Doppelmembran mit Poren umgeben. Diese Poren haben einen Durchmesser von 40 bis 100 nm und lassen Makromoleküle passieren. Die Kerndoppelmembran hat direkte Verbindung zum Endoplasmatischen Reticulum (ER).
-
Schematischer Bau eines Eucyten
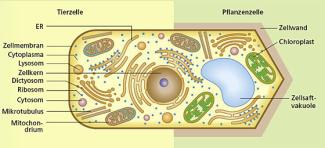
Renate Diener

Im Kerninneren befindet sich das Karyoplasma mit Cytoskelett-Strukturen (Kernskelett). Mit geeigneter Färbetechnik lässt sich im Kern ein nucleinsäurehaltiges Netzwerk, das Chromatin, nachweisen. Es besteht aus den teilweise entspiralisierten Chromosomen und enthält den größten Teil der Erbinformation einer Zelle. Die Nucleotidsequenz der DNA und die Anzahl der Chromosomen sind artspezifisch. Im Zellkern findet man außerdem meist zwei Kernkörperchen (Nucleoli). Sie bestehen vorwiegend aus RNA und wirken bei der Bildung der Ribosomen mit.
Während der Mitose und Meiose löst sich die Kernmembran auf. Die Chromosomen werden sichtbar. Nach der Ausbildung der Chromatiden und deren Verteilung auf die beiden entstehenden Zellen bzw. der Trennung der homologen Chromosomen und deren Verlagerung in die Tochterzellen bildet sich die Kernmembran vom ER aus wieder neu.
Wie der Zellkern so sind auch die 1 bis 10 nm langen Mitochondrien Organellen mit zwei Membranen als Hülle. Die innere Membran ist vielfach gefaltet und eingestülpt. An dieser großen Oberfläche können zahlreiche Stoffwechselvorgänge gleichzeitig ablaufen. Durch die Doppelmembran entstehen zwei Kompartimente: Der Intermembranraum (perimitochondrialer Raum) und die Matrix. Die Außenmembran gilt als relativ durchlässig. Durch die Innenmembran gelangen Stoffe nur mithilfe von Transportproteinen. Sie ist der Sitz der Enzyme für die Atmungskette und der ATP-Synthese.
Die Anzahl der Mitochondrien in einer Zelle hängt sehr von deren physiologischer Aktivität ab. Während Zellen der Säugerleber bis zu 1 500 Mitochondrien enthalten, sind es in manchen Eizellen etwa 100 000 und in einzelligen Amöben um die 50 000.
Plastiden sind typische Organellen von Algen und Pflanzen. Sie können sehr unterschiedlich gestaltet sein. Plastiden vermehren sich durch Teilung über ein als Proplasmid bezeichnetes Vorstadium. Als Chloroplasten ermöglichen sie ihren Trägern eine autotrophe Lebensweise. Fotosynthetisch aktive Zellen höherer Pflanzen besitzen durchschnittlich 50 bis 100 Chloroplasten von linsenförmiger Gestalt und etwa 5 µm Größe. Besonders in grünen Algen kommen auch anders geformte Chloroplasten und manchmal in sehr geringer Anzahl vor.
Die bereits lichtmikroskopisch sichtbaren Chloroplasten sind von einer Doppelmembran umgeben. Die Außenmembran grenzt sie gegen das Cytoplasma ab.
Von der Innenmembran geht ein weitverzweigtes, geordnetes Membransystem, das Thylakoidsystem , aus. Dieses Thylakoidsystem lässt sich in zwei Bereiche gliedern: Die Granathylakoide und die Stromathylakoide.
Die Granathylakoide bilden geordnete Stapel von Doppelmembranen. Diese als dunkelgrüne Areale im Chloroplasten erscheinenden Lamellenstapel enthalten reichlich Fotosynthesepigmente und alle für die Lichtreaktion erforderlichen Enzyme. Die Stromathylakoide stellen Verbindungen zwischen den Grana her. Sie bilden keine Stapel aus, sondern durchziehen das Plastideninnere in Form von Membranflächen. Das Thylakoidsystem ist in die Grundsubstanz des Chloroplasten, das Stroma oder die Matrix, eingebettet. Im Stroma befinden sich auch die Enzyme für die lichtunabhängige Reaktion. Während der Alterung, etwa im Zuge der Herbstfärbung, degeneriert das Membransystem bei gleichzeitigem Abbau des Chlorophylls.
-
Bau eines Chloroplasten
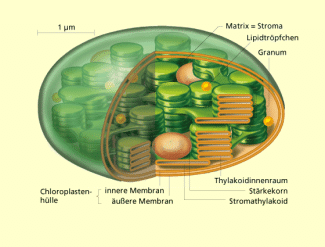
Renate Diener

Mitochondrien und Plastiden sind aus endosymbiontischen Prokaryoten hervorgegangen und sie verfügen bis heute über eine eigene DNA (mt-DNA bzw. p-DNA). Da sie auch Ribosomen des 70 S-Typs enthalten, sind sie in der Lage, einige Enzyme selbst herzustellen. Sie vermehren sich durch einfache Teilung.
Das Endomembransystem der Zelle besteht aus Endoplasmatischem Reticulum, Golgi-Apparat (Dictyosomen), Lysosomen und Cytosomen (Microbodies). Das Endoplasmatische Reticulum (ER) durchzieht in ausgewachsenen Zellen als reich gegliedertes Membransystem das Cytoplasma. Es steht mit verschiedenen Zellorganellen, etwa dem Kern und den Dictyosomen, in enger Verbindung und kompartimentiert das Zellinnere. Die Zisternen genannten Hohlräume zwischen den beiden Membranen sind intrazelluläre Speicherorte und Transportkanäle, an deren Ende Vesikel abgeschnürt werden. Die Außenseiten der ER-Membranen können ohne oder mit Ribosomen besetzt sein. Entsprechend unterscheidet man glattes (agranuläres) von rauem (granulärem) ER. Das raue ER ist der Syntheseort zahlreicher Proteine, vor allem der Speicher- und Membranproteine. Das glatte ER wirkt bei sehr unterschiedlichen Stoffwechselvorgängen mit.
-
Bau des Endoplasmatischen Reticulums
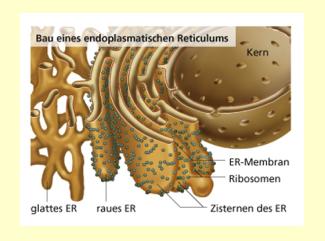
Renate Diener

Dictyosomen oder der Golgi-Apparat als die Gesamtheit der Dictyosomen in einer Zelle können als Membranstapel mit besonders aktiven Zisternen aufgefasst werden. Die Bezeichnung Golgi-Apparat erfolgte nach seinem Entdecker CAMILLO GOLGI, der 1898 das plasmatische Zisternensystem in der Zelle fand. An Dictyosomen sammeln sich Polysaccharide oder Proteine, die nach Vesikelabschnürung in andere Zellorganellen gelangen oder als „coated vesicle“ die Zelle verlassen. In Zellen mit sekretorischer Funktion ist der Golgi-Apparat besonders stark ausgeprägt. In Pflanzenzellen produzieren Dictyosomen beispielsweise Pectine und Hemicellulosen, in tierischen Zellen Verdauungsenzyme und Hyaluronsäure.
Lysosomen sind von einer einfachen Membran umgeben und enthalten Enzyme. Gebildet werden diese Organellen vom Golgi-Apparat. Aufgabe der Lysosomen in der Zelle ist es, sehr selektiv mit Endocytosevesikeln zu verschmelzen und den Vesikelinhalt hydrolytisch abzubauen. Beim Absterben von Zellen löst sich die Lysosomenmembran auf. Die Enzyme werden frei und autolysieren die Zelle. Bislang sind mehr als 60 verschiedene lysosomale Enzyme nachgewiesen worden. Als Leitenzym gilt die saure Phosphatase. Infolge von Defekten an Lysosomen können beim Menschen Erbkrankheiten auftreten. Mehr als 30 derartiger Krankheiten sind bekannt.
0,5 bis 1,5 µm große, von einer Membran umgebene und mit bestimmten Enzymen gefüllte Vesikel bezeichnet man als Microbodies (Cytosomen). Cytosomen schnüren sich, im Gegensatz zu den Lysosomen, nicht vom Endomembransystem ab, sondern gehen aus bereits bestehenden durch Teilung hervor. Der Inhalt dieser kleinen Zellorganellen wird von freien, cytoplasmatischen Ribosomen synthetisiert. Bisher sind nahezu 50 verschiedene Enzyme in den Microbodies nachgewiesen worden. Entsprechend dem Inhalt differenziert man nach Peroxisomen (u. a. Oxidasen, Katalasen) in tierischen Zellen und Blatt-Peroxisomen und Glyoxisomen (u. a. Isocitrat-Lyase, Malat-Synthase).
Sowohl bei tierischen als auch bei pflanzlichen Zellen treten Vakuolen auf. In tierischen Zellen sind es nur kleine, mit Flüssigkeit angefüllte Räume. In älteren Pflanzenzellen erreichen sie ein beachtliches Volumen. In manchen Zellen entsteht aus mehreren einzelnen Vakuolen eine mächtige Zentralvakuole. Die einfache Membran, die den Vakuoleninhalt zum Cytoplasma hin abgrenzt, wird als Tonoplast bezeichnet. Dieser Tonoplast unterscheidet sich zwar nicht im grundsätzlichen Aufbau, jedoch in den Membraneigenschaften deutlich vom Plasmalemma. So sind Plasmalemma und Tonoplast für verschiedene Stoffe unterschiedlich durchlässig.
Vakuolen können osmotisch Wasser aufnehmen und abgeben. Der davon abhängige Binnendruck (Turgor) einer Pflanzenzelle hat Bedeutung für die Stabilität des Pflanzengewebes. Heute weiß man, dass sie in pflanzlichen Zellen mehrere wichtige Funktionen erfüllen. Sie können den Lysosomen gleich lytische Enzyme enthalten und so bestimmte Stoffumsetzungen beeinflussen. Vakuolen dienen aber auch als Deponierraum für überflüssige, oft toxische Sekundärstoffe. Sie ersetzen und ergänzen die ansonsten spärlichen Ausscheidungsmöglichkeiten der Pflanze. Der Vakuoleninhalt besteht dann vor allem aus Glykosiden, Phenolen oder Pigmenten. Bedeutsam können Vakuolen auch für die Speicherung von Stoffen sein.
Sowohl verschiedene Zucker als auch Proteine und Fette erreichen in den Vakuolen zum Teil beachtliche Konzentrationen. Sie sind somit zunächst dem aktuellen Stoffwechselgeschehen entzogen. Bei Bedarf werden diese Reservestoffe wieder mobilisiert. Vakuolen spielen auch eine entscheidende Rolle bei der Aufrechterhaltung der Turgeszenz von Zellen. Der Zellsaft enthält gelöste, osmotisch wirksame Verbindungen. Sie machen den osmotischen Wert des Vakuoleninhalts aus. Dieser bestimmt im Wesentlichen die Intensität und Richtung des Wasserstroms in die oder aus der Zelle.
Ribosomen sind die Orte der Proteinbiosynthese in der Zelle. Sie stellen Organellen ohne Membranhülle dar. Zwei Ribosomen-Grundtypen werden unterschieden: die 80 S-Ribosomen im Cytoplasma der Eucyte und die vom 70 S-Typ der Plastiden, Mitochondrien und der Procyte. Gemeinsam ist beiden Typen, dass das komplette Ribosom aus zwei Untereinheiten aufgebaut ist. 70 S-Ribosomen setzen sich aus 30 S- und 50 S-Untereinheiten zusammen, 80 S-Ribosomen aus 40 S- und 60 S-Untereinheiten. Ribosomen und ihre Untereinheiten werden entsprechend ihrem Sedimentationskoeffizienten bei der Ultrazentrifugation bzw. Dichtegradientenzentrifugation nach SVEDBERG-Einheiten (S) benannt.
Der Wert einer größeren Einheit (80 S) ergibt sich nicht aus der Summe der Werte von Untereinheiten. (Aufgrund der unterschiedlichen Formen lassen sich keine nummerischen Summen aus den Untereinheiten zum Gesamtribosom bilden – 30 S- und 50 S-Untereinheiten ergeben also nicht 80 S, sondern der Koeffizient des ganzen Ribosoms liegt bei 70 S; ebenso ergibt sich bei 40 S- und 60 S-Untereinheiten nicht die Summe 100 S, sondern der Sedimentationskoeffizient des eukaryotischen Gesamtribosoms liegt bei 80 S.)
Während der Proteinbiosynthese (Translation) reihen sich 5 bis 40 Monosomen an der mRNA zu einer Polysomenkette auf.
In Procyten hat man um die 10 000 Ribosomen pro Zelle, in stoffwechselaktiven Säugerzellen ein Vielfaches davon gefunden. Die Lebensdauer der Ribosomen kann relativ kurz sein. Deshalb müssen sie innerhalb eines Ribosomenzyklus ständig neu gebildet werden. Bis zu 100 Ribosomen pro Sekunde können es in einer Wirbeltierzelle sein. Funktionsfähige Ribosomen bestehen aus Proteinen und RNA in einem Mengenverhältnis von 1 : 1. Im Zellkern wird die ribosomale RNA synthetisiert und mit Proteinen beladen. Als Vorstufen der Ribosomen-Untereinheiten passieren sogenannte Präribosomen die Poren der Kernmembran und gelangen so in das Cytoplasma. Dort erhalten sie ihre endgültige Ausprägung.
Das Vorhandensein von zwei Ribosomentypen in der gleichen Zelle gilt als wesentliche Stütze der Endosymbiontentheorie. Ursprünglich eigenständige Vorläufer der Plastiden und Mitochondrien haben die 70 S-Ribosomen ebenso wie eine eigene DNA bei der Cytosymbiose mit eingebracht. Allerdings können die 70 S-Ribosomen heute nur noch einen kleinen Teil der Proteine vollständig selbst herstellen. Die Mehrzahl der für die Organellen erforderlichen Proteine wird an den cytoplasmatischen Ribosomen oder in Kooperation mit ihnen synthetisiert.
Nicht in allen eukaryotischen Zellen kommen Centriolen vor. Während sie in tierischen Zellen stets paarweise auftreten, fehlen sie den Eucyten der Bedecktsamer (Angiospermen). Centriolen sind von tubulärer, geordneter Struktur. Sie gelten allgemein als Organisationszentren für Mikrotubuli und spielen eine wichtige Rolle bei der Ausbildung von Basalkörpern der Geißeln und beim Aufbau der Kernteilungsspindel während der Mitose und Meiose.
Die Zellwand umgibt die pflanzliche Eucyte mit einer festen, Form gebenden Hülle. Während des Lebens einer Zelle durchläuft die Wandbildung verschiedene Phasen. Nach erfolgter Kernteilung entsteht zwischen den beiden Tochterzellen von innen heraus (zentrifugal) eine neue Trennschicht. Protopectin- und Hemicellulose-Moleküle ergeben eine Mittellamelle. Während des folgenden Zellstreckungswachstums lagern sich zunächst ungeordnet beidseitig Mikrofibrillen aus Cellulose ein. Die Primärwand entsteht. Sobald das Streckungswachstum abgeschlossen ist, verstärkt sich die Wand durch schichtweise Auflagerung weiterer Cellulosefibrillen (Sekundärwand). Ein- und Auflagerungen von Wachs, Lignin und Suberin imprägnieren die Wand und verleihen ihr Festigkeit und Widerstandsfähigkeit (Tertiärwand).
Pflanzliche und tierische Zellen können über Plasmabrücken (Plasmodesmen) in Verbindung stehen. Die Pflanzenzelle unterscheidet sich von den tierischen Eucyten durch den Besitz von Plastiden, einer großen Zentralvakuole und einer Zellwand. Im Vergleich zu pflanzlichen Eucyten bilden tierische Zellen häufig Fortsätze aus. Ein charakteristisches Merkmal tierischer Zellen ist auch der Besitz von zwei Centriolen.
-
Schematischer Aufbau der Zellwand einer ausgewachsenen Pflanzenzelle
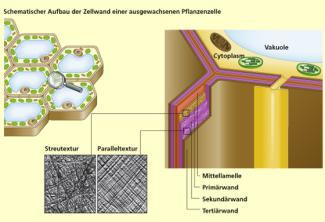
Walther-Maria Scheid

Suche nach passenden Schlagwörtern
- Turgor
- Euzyten
- Sekundärwand
- Stromathylakoide
- Kernkörperchen
- Mikrotubuli
- Dictyosomen
- ER
- Primärwand
- Microbodies
- Thylakoidmembran
- Speicherung von Stoffen
- Chromatin
- Cytosol
- Zellkern
- Tonoplast
- Zellen
- Plastiden
- Golgi-Apparat
- Plasmalemma
- Lysosomen
- Zellwand
- Ribosomenzyklus
- Endoplasmatische Reticulum
- Vakuolen
- Endosymbiontentheorie
- Video
- Chloroplasten
- Plasmabrücken
- Matrix
- Endomembransystem
- Cytoskelett
- Procyten
- Nucleus
- Mitochondrien
- Camillo Golgi
- Plasmolyse
- Granathylakoide
- Cytoplasma
- Cytosomen
- Plasmodesmen
- Eukaryoten
- Tertiärwand
- Intermembranraum
- Eucyten
- Peroxisomen
- Prozyten
- Karyon
- Thylakoidsystem
- Centriolen
- Ribosomen
- Turgeszenz

