Inselökologie
Die Artenzahl auf Inseln wird von ihrer Größe, ihrer Entfernung zu Festländern bzw. zu anderen Inseln, ihrer geografischen Lage – insbesondere der geografischen Breite – und ihrer Habitatvielfalt bestimmt. Nach dem Gleichgewichtsmodell von ROBERT MACARTHUR und EDWARD OSBORNE WILSON (1967) stellt sich auf Inseln unter Normalverhältnissen ein Gleichgewicht zwischen Einwanderungsrate und Aussterberate ein.
Da die Evolution auf einer Insel anders verläuft als auf benachbarten Festländern oder auf anderen Inseln, beherbergen Inseln je nach ihrer Isolation einen mehr oder weniger großen Prozentsatz endemischer (nur auf dieser Insel vorkommender) Arten. Einmal können dies Reliktendemiten sein, nämlich Lebewesen, die ursprünglich eine viel weitere Verbreitung hatten und die mangels Konkurrenz auf dieser Insel überleben konnten – wie etwa die Brückenechse auf einigen kleinen Inseln nördlich von Neuseeland – zum anderen können sich eingewanderte Arten isoliert von ihrer Stammart zu neuen Arten weiterentwickeln. Durch Anpassung an unterschiedliche Habitate (Einnischung) können so aus einer Stammart durch adaptive Radiation (rasche Folge von Artaufspaltungen, ausgehend von einem gemeinsamen Vorfahren) viele Tochterarten entstehen (Beispiel: die Darwinfinken auf Galapagos, Fruchtfliegenarten auf Hawai).
Inseln isolieren
Das Wort „Isolation“ lässt sich – ebenso wie das Wort „Insel“ – auf das lateinische „insula“ zurückführen, heißt also eigentlich „Verinselung“.
Seit langem weiß man, dass Inseln weniger Arten beherbergen als vergleichbar große Festlandsgebiete. Natürlich hängt die biologische Vielfalt, also die Artenzahl einer Insel, von ihrer Fläche ab. Dies ist sogar eine verhältnismäßig einfache Beziehung, die sich aus empirischen Daten ergibt und die als Artenzahl gegen den Logarithmus der Inselfläche aufgetragen meist eine mehr oder weniger deutliche Gerade ergibt. In arithmetischer Darstellung ergibt sich immer eine mehr oder weniger gekrümmte Kurve, deren Steigung mit zunehmender Fläche nahezu gegen Null gehen kann. Natürlich gibt es viele weitere Parameter, die für die Artenvielfalt einer Insel verantwortlich sind, z. B. die Vielfalt der Habitate (auch die ist in der Regel wiederum flächenabhängig), die Entfernung von einem großen Festland oder von anderen Inseln, das Alter, die geografische Breite usw.
-
Artenzahl-Arealkurve der Vögel auf den Salomonen und weiterer Beispiele. Links sind die Beziehungen für Inselarten logarithmisch, rechts arithmetisch dargestellt. a) Vögel auf den Salomonen, b) Amphibien und Reptilien auf den Westindischen Inseln, c) Fische in den unterschiedlichen Quelltöpfen australischer Wüstengebiete, d) Wirbellose Tiere in Muschelbänken
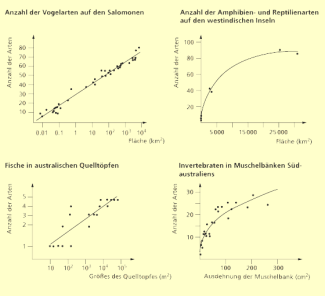

„Insel“ bedeutet in diesem Zusammenhang übrigens nicht unbedingt eine allseits von Wasser umgebene Landfläche – so werden Inseln in Lexika im Allgemeinen definiert. Auch Seen sind Inseln in einem Meer von Land, Bergspitzen in einer Ebene, Lichtungen in einem Wald, Oasen in einer Wüste. Ebenso gibt es Inseln, die sich in geologischen Merkmalen, Bodentypus oder Vegetation von ihrer Umgebung unterscheiden. Auch Verkehrsinseln sind Inseln in einer biologisch feindlichen Straßenumgebung. Für diese Formen von Inseln ergeben sich sehr ähnliche Kurven, wenn Artenzahl und Arealgröße gegeneinander aufgetragen werden. So schreiben schon ROBERT MACARTHUR (1930–1972) und EDWARD OSBORNE WILSON (*1929) 1971, dass „die inselartige Beschaffenheit ein überall anzutreffendes Merkmal der Biogeografie ist“.
Daraus wird deutlich, dass Fragen der Inselökologie oder Inselbiogeografie weit über die Bedeutung eigentlicher Inseln hinausgehen, wenn man die Verteilung und das Muster von Arten und Artengemeinschaften untersuchen will. Denn es gibt kaum natürliche Lebensgemeinschaften, die nicht zumindest einige Elemente von Inselhaftigkeit besitzen. Schon die Unterteilung der Biosphäre in verschiedene Ökosysteme zeigt ja mit dem Begriff „Ökosystem“, dass man sich die Biosphäre zusammengesetzt vorstellt aus Teilsystemen, die voneinander mehr oder weniger isoliert, und damit verinselt, sind.
Einwanderung – Auswanderung
Nach der Theorie der Habitatdiversität, die v. a. auf D. LACK (Island Biology Illustrated by the Land Birds of Jamaica, 1976) zurückgeht, ist die Artenzahl einer Insel in erster Linie von der Zahl der unterschiedlichen Lebensräume abhängig. Da diese Zahl der Lebensräume in der Regel mit der Flächengröße in Beziehung steht, kommt es nach LACK zu den vorher erwähnten Artenzahl-Arealkurven. Zwingend ist dies jedoch nicht, habitatarme, größere Inseln können artenärmer sein als habitatreiche, kleinere Inseln. Da sich LACK nur mit Vogelarten beschäftigte, ist auch verständlich, dass er sich mit Zuwanderungsbarrieren weniger befasste. Nach seiner Vorstellung beruht die Abwesenheit bestimmter Populationen auf einer Insel nicht auf mangelnder Besiedelung, sondern immer auf dem Fehlen geeigneter Lebensräume (Habitate).
Demgegenüber begründeten MACARTHUR und EDWARD O. WILSON 1967 die Gleichgewichtstheorie der Inselbesiedelung. Danach stellt sich – qualitativ leicht zu beschreiben – auf jeder Insel ein Gleichgewicht zwischen Einwanderungsrate und Aussterberate der Arten ein. Je mehr Arten auf einer Insel vorhanden sind, desto geringer ist die Einwanderungsrate. Entweder, da keine Arten zur Einwanderung mehr zur Verfügung stehen oder da es keinen Platz mehr für die neu zugekommenen Arten gibt, da keine „Nische“ mehr für sie frei ist. Umgekehrt ist die Aussterberate umso größer, je mehr Arten auf der Insel sind. Im einfachsten Fall könnte es sich hier um eine lineare Beziehung handeln. Dann ließe sich dies in einem Raten-Artenzahldiagramm mit Geraden darstellen. Der Schnittpunkt wäre der Gleichgewichtspunkt, in dem sich der Artenreichtum im Gleichgewicht befindet. Wahrscheinlicher ist allerdings, dass es sich bei der Beziehung von Aussterberate bzw. Einwanderungsrate zu Artenzahl nicht um eine lineare Beziehung handelt, sondern eher um zwei konkave Kurven wie in der Abbildung dargestellt.
Verschiedene Diagramme zur Gleichgewichtstheorie von MacArthur und Wilson: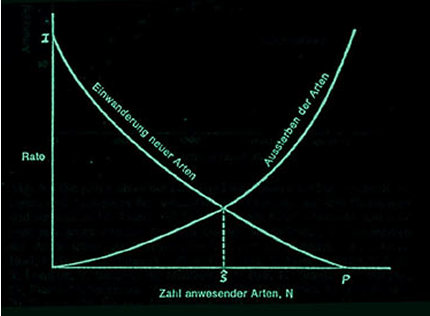 Im einfachsten Fall ergibt sich eine lineare Beziehung zwischen Einwanderungsrate und Artenzahl sowie zwischen Auswanderungsrate und Artenzahl. Der Schnittpunkt beider Geraden gibt die Gleichgewichtsartenzahl auf der Insel an.
Im einfachsten Fall ergibt sich eine lineare Beziehung zwischen Einwanderungsrate und Artenzahl sowie zwischen Auswanderungsrate und Artenzahl. Der Schnittpunkt beider Geraden gibt die Gleichgewichtsartenzahl auf der Insel an.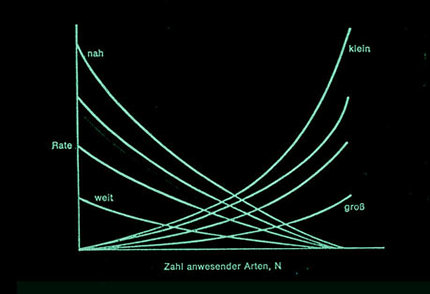 Einwanderungsrate und Aussterberate hängen vom Abstand der Insel von einem größeren Festland und von der Inselgröße ab. Große und nahegelegene Inseln beherbergen mehr Arten als kleine, entfernt gelegene.
Einwanderungsrate und Aussterberate hängen vom Abstand der Insel von einem größeren Festland und von der Inselgröße ab. Große und nahegelegene Inseln beherbergen mehr Arten als kleine, entfernt gelegene.
In jedem Fall stellt sich – steht nur genügend Zeit zur Verfügung – ein Gleichgewicht ein, eine bestimmte Artenanzahl. Die Zusammensetzung der Arten, das Artenspektrum, kann oder muss sich allerdings weiter ändern, da ja immer Arten aussterben und Arten einwandern, in einer Rate, die dem Gleichgewicht entspricht.
Dabei hängt der Kurvenverlauf sehr stark von der Beschaffenheit der Insel und ihrer Lage zu einem benachbarten Festland ab. Bei nahe zum Festland gelegenen Inseln ist die Einwanderungsrate zunächst sehr groß, bei weit entfernten klein. Auch die Größe der Insel spielt eine Rolle. Je größer, desto mehr Einwanderer können „aufgefangen“ werden. Dabei spielt auch die Gestalt der Insel eine Rolle.
Umgekehrt ist die Aussterberate bei großen Inseln geringer als bei kleinen, da sie viel mehr Lebensräume enthalten und deshalb auch Platz für mehr ökologische Nischen sprich Arten bieten.
Man kann eine Schar unterschiedlicher Kurven für Einwanderungsrate und Aussterberate in Abhängigkeit von der Artenzahl auftragen und erhält dann – je nach Kombination – unterschiedliche Gleichgewichtswerte. Auch die Entfernung von Nachbarinseln spielt für die Einwanderungsraten eine wichtige Rolle: Die Artenzahl eng benachbarter Inseln ist größer als die Artenzahl sonst identischer Inseln, die weiter voneinander entfernt liegen.
Obwohl dies zunächst sehr plausibel klingt, so zeigte sich doch bei der empirischen Überprüfung, dass es ausgesprochen schwierig ist, dieses Modell durch eindeutige Daten zu belegen. So lässt sich die Größenabhängigkeit der Artenzahl im Gleichgewicht auch alleine durch Habitatdiversität erklären. Und auch der Einfluss der Abgelegenheit kann vollkommen unabhängig von der Gleichgewichtstheorie betrachtet werden. Viele Arten sind nämlich in ihrer Ausbreitung so limitiert, dass sie entfernte Inseln nicht nur erschwert erreichen, sondern auch erst sehr viel später. Dies kann dazu führen, dass solche isolierten Inseln weniger „gesättigt“ sind, dass sich also das von MACARTHUR und WILSON beschriebene Gleichgewicht noch gar nicht eingestellt hat. In jedem Fall sollte man die Theorie der Habitatvielfalt, also die Aussage, dass die Vielfalt der Lebensräume für die Artenzahl auf einer Insel verantwortlich ist (LACK), und die Aussage, dass dafür das sich einstellende Gleichgewicht aus Einwanderungsrate und Aussterberate verantwortlich ist (MACARTHUR und WILSON) nicht als gegensätzliche, sondern als sich gegenseitig ergänzende Theorien ansehen.
Zyklen
Die beiden oben erwähnten Modelle zur Inselbesiedelung gehen von einem Endgleichgewichtszustand aus, auch wenn es sich bei dem Einwanderungs-Aussterbe-Modell eher um ein „Fließgleichgewicht“ handelt. Die Entwicklung strebt einem ausgeglichenen Endzustand zu. Einen solchen Zustand nennt man in der Biologie einen Klimaxzustand. An diesem „biologischen Gleichgewicht“ verändert sich dann nichts mehr, es sei denn, es kommt durch Eingriffe von außen oder durch Katastrophen dazu, dass alles wieder von vorne anfängt. Nach der Klimaxvorstellung sind das aber Ausnahmen.
Dem steht die dynamische Auffassung über Biozönosen gegenüber, die am besten durch die Mosaik-Zyklus-Theorie (H. REMMERT, W. SCHERZINGER 1991) beschrieben wird. Am deutlichsten wird dies bei der Untersuchung von (Ur-)Waldökosystemen. So gibt es z. B. keinen Endzustand eines natürlichen Urwalds. Vielmehr entwickeln sich nach dem Zusammenbrechen der alten Bäume Lichtungen und dort entstehen unter heftiger Konkurrenz zunächst Gesellschaften von Pionierbaumarten, die ihrerseits nach einiger Zeit wieder zusammenbrechen. Auch dann kommt es wieder zu heftiger Konkurrenz und schließlich kehren die ursprünglichen Baumarten zurück. In einem großen Waldökosystem laufen diese Prozesse ständig auf kleinen Flächen („patches“) nebeneinander ab, sodass immer viele verschiedene Entwicklungsstadien mosaikartig nebeneinander liegen. Ein besonders schönes Beispiel für diese, sich im Kreis entwickelnde Dynamik zeigt die Abfolge der Waldstadien in einer flachen Senke, die durch die Aktivitäten von Bibern zu einem See aufgestaut wurde. Es kommt z. B. durch Blaugrüne Bakterien zu einer ganz erheblichen Stickstoffanreicherung in diesem See. Der See verlandet. Auf dem mineralstoffreichen humosen Boden entwickelt sich zunächst eine üppige Staudenvegetation stickstoffliebender Pflanzen, dann siedeln sich erste Weichhölzer an und ganz zum Schluss kommt es zu einer Wiederbesiedelung durch die ursprünglichen Waldbaumarten.
In Wäldern können solche Zyklen Jahrhunderte, vielleicht sogar Jahrtausende dauern, in anderen Vegetationsformen nur Jahrzehnte.
Natürlich gelten diese Überlegungen auch für Inseln, auch hier sind Veränderungen und kleine Katastrophen eher die Regel als die Ausnahme. Dies müssen keine ständigen Vulkanausbrüche sein (wie etwa auf Hawaii), auch epedemieartig auftretende Krankheiten an dominierenden Baumarten können ganz neue Zyklen einleiten.
„Ein sehr großer Teil der auf der Roten Liste stehenden Pflanzen- und Tierarten unserer Heimat sind an derartige Sukzessionsstadien gebunden“ (REMMERT 1984).
Wenn es darum geht, Biodiversität zu schützen und zu erhalten, können solche eher theoretischen Überlegungen wichtige Voraussetzungen für Planung und Eingriffsoptimierung sein.
Inselökologie und Naturschutz
„Die Landschaften Mitteleuropas weisen eine wachsende Tendenz der Verinselung der einzelnen sie bildenden Landschaftsbestandteile auf. Die Isolationswirkung zwischen den teilweise nur noch als Fragmenten erhaltenen Resten ursprünglicher Landschaftselemente nimmt zu. Damit verliert die Landschaft auch funktional die Eigenschaft eines vielfach engmaschig verbundenen Netzes und entwickelt sich stattdessen zu einem komplex mosaikartigen Nebeneinander existierender Teilstücke.“ (H. J. MADER 1984). Um biologische Vielfalt zu erhalten ist es deshalb wichtig, Barrieren abzubauen, wertvolle Biotope und Schutzgebiete miteinander zu vernetzen und Trittsteine einzubauen. Ausgeräumte Agrarlandschaften können durch Wallhecken („Knicks“) und Feldgehölze erheblich an Artenvielfalt gewinnen, Stadtbrachen und ringförmig und radiär verlaufende Grünzüge in Städten können die Biodiversität erhöhen. Ein besonderes Problem stellt in dichtbesiedelten Räumen die Zerschneidung der Landschaft durch Verkehrswege, insbesondere durch Straßen, dar. Dieser Zerschneidungseffekt kann z. B. durch die Einrichtung von Biotopbrücken über Autobahnen erheblich gemindert werden.
Inselendemiten
Arten sind keine unveränderlichen konstanten Einheiten. Sie verändern sich, sie können sich in neue Arten aufspalten. Damit können auf Inseln ganz neue Lebensgemeinschaften entstehen, deren Vorkommen nur sehr indirekt mit Einwanderung und Aussterben begründet werden kann. Auf der nur 12 qkm großen Karibikinsel Utila 50 km vor der Küste von Honduras wurde im 20. Jahrhundert eine Leguanart – Ctenosaurus bakeri – entdeckt, die nur dort vorkommt. Naturschutzinitiativen bemühen sich um die Erhaltung dieser nur noch auf 3 000 Individuen geschätzten Reptilienart. Solche Arten, die nur ein eng begrenztes Areal besiedeln, nennt man Endemiten. Endemiten sind typisch für Inseln. Endemiten der Kanarischen Inseln z. B. sind die Dickblattgewächse der Gattung Aeonium, die dort und nur dort mit über 40 Arten vorkommen, von denen viele auch noch auf einzelne Inseln beschränkt sind. Ein Kanarenendemit ist auch die rötlich blühende und als Liane wachsende Kanaren-Glockenblume (Canaria canariensis) oder der große Kanaren-Natternkopf Echium wildpretii. Bei manchen Arten – wie etwa beim Tëide-Finken – kann man die Verwandtschaft zu weitverbreiteten Festlandsarten durchaus erkennen. Der Tëide-Fink sieht aus wie ein blaustichiges Foto eines Buchfinken. Im Allgemeinen ist der Anteil der Endemiten an dem Artenbestand einer Insel umso größer, je länger die Insel besteht, auch je stärker sie isoliert ist. Die absolute Anzahl hängt natürlich auch wieder von der Inselgröße und ihrer Habitatvielfalt ab.
Besonders berühmt durch die Vielzahl ihrer Inselendemiten wurde das Galapagos-Archipel – nicht zuletzt, weil CHARLES DARWIN (1809–1882) diese einsame Inselgruppe auf seiner Weltreise 1834 besuchte und dort wichtige Impulse für die Entwicklung seiner Evolutionstheorie erhielt. Ihm folgten und folgen bis heute viele Biologen und auch naturkundlich interessierte Touristen.
Galapagosendemiten, von denen immer wieder die Rede ist, sind z. B. die Erdfinken (Gattung Geospiza), die auch „Darwinfinken“ genannt werden oder die Riesenschildkröten der Art Chelonoidis elephantopus, die mit 11 Unterarten ebenfalls nur auf den Galapagos Inseln zu finden sind. DARWIN berichtet in seinem Reisebericht „Reise eines Naturforschers um die Welt“ ausführlich von den „Elefantenschildkröten“. Unter anderem schreibt er:
„Noch habe ich den allermerkwürdigsten Zug der Naturgeschichte dieses Archipels nicht erwähnt; er besteht darin, dass von den verschiedenen Inseln in beträchtlichem Maße jede von einer verschiedenen Gruppe von Geschöpfen bewohnt wird. Meine Aufmerksamkeit wurde zuerst dadurch auf diese Tatsache gelenkt, dass der Vizegouverneur Lawson erklärte, die Schildkröten von den verschiedenen Inseln seien untereinander verschieden, und er könne mit Sicherheit sagen, von welcher Insel irgendeine hergebracht sei. Eine Zeitlang schenkte ich dieser Angabe nicht hinreichende Aufmerksamkeit und ich hatte bereits zum Teil die Sammlungen von zwei der Inseln untereinander gemengt. Es wäre mir doch nicht im Traume eingefallen, dass ungefähr fünfzig oder sechzig Meilen voneinander entfernt liegende Inseln, die meisten in Sichtweite voneinander, aus genau denselben Gesteinen bestehend, in einem ganz gleichartigen Klima gelegen und nahezu zu derselben Höhe sich erhebend, verschiedene Bewohner haben sollten; wir werden aber sofort sehen, dass dies der Fall ist. Es ist das Geschick der meisten Reisenden, sobald sie entdeckt haben, was an irgendeinem Ort das Interessanteste ist, eiligst fortgetrieben zu werden; ich muss aber gerade dafür dankbar sein, dass ich genügendes Material erhalten konnte, diese äußerst merkwürdige Tatsache in der Verbreitung der organischen Geschöpfe ermitteln zu können.“
IRENÄUS VON EIBL-EIBESFELDT (*1928) schreibt in seinem Klassiker „Galapagos“ über diese Echsen:
„Diese Elefantenschildkröten sind Überreste einer einst weit über die Erde verbreiteten Tiergruppe. Die Ordnung der Schildkröten hat sich seit dem Erdmittelalter nur wenig geändert. Triassochelys dux aus dem Keuper von Halberstadt hat bereits den typischen Knochenpanzer, der den Körper wie eine Kapsel umschließt. Gewaltige Landschildkröten waren vor 60 Millionen Jahren in Europa, Amerika und Indien beheimatet. Nach einigen Überresten zu schließen, wogen manche Exemplare über eine Tonne. Mit dem Auftreten der wendigen Säuger, die wohl vor allem den Jungen und Eiern nachstellten, wie das heute auf Galapagos die eingeschleppten Säuger tun, verschwanden die Riesenschildkröten in den meisten Erdgebieten. Sie hielten sich nur auf einigen ursprünglich von Säugern freien Inseln, nämlich auf den Maskarenen im Indischen Ozean und auf den Galapagos-Inseln. Mensch und Haustiere haben diese Bestände dezimiert. Die Maskarenen-Schildkröte konnte sich nur auf Aldabra halten.“
Auch für die Endemiten gilt – wie für die Überlegungen zur Biodiversität – dass man nicht nur bei wirklichen Inseln, also von Wasser umgebenen Festländern, sondern auch bei allen anderen inselartigen Lebensräumen ähnliche Verhältnisse vorfindet. Besonders berühmt sind die endemischen Buntbarsche alter afrikanischer Seen wie etwa des Malawi-Sees oder des Tanganjika-Sees. Im Tanganjika-See wurden 214 Buntbarsch-Arten nachgewiesen, von denen 80 % nur in diesem See vorkommen. Dabei zeigen gerade die afrikanischen Seen sehr gut, dass die Zahl der Endemiten etwas zu tun hat mit dem Alter der „Insel“. Der Tanganjika-See wird auf ein Alter von 12 Millionen Jahren geschätzt. Der viel jüngere Rudolph-See dürfte erst seit 5 .000 Jahren vom Flusssystem des oberen Nils getrennt sein. In dieser Zeit konnten sich in diesem See immerhin fünf endemische Cichliden-Arten entwickeln.
Vielfalt und Evolution
„Das größte Wunder unseres Planeten ist die ungeheure Vielfalt der Lebensformen. So zahlreich sind die Arten, dass wir die meisten von ihnen noch gar nicht identifiziert haben. Die Biosphäre bedeckt die Erde mit einem Teppich aus kunstvoll miteinander gekoppelten Lebensformen. Sogar die scheinbar öde arktische Tundra beherbergt viele Tier- und Pflanzenarten einschließlich der mannigfaltigen Gruppe symbiotischer Flechten, die untereinander und mit ihrer Umwelt ein kompliziertes Netz von Wechselbeziehungen aufrechterhalten.“
Dies sind die ersten Sätze aus dem Vorwort des von EDWARD O. WILSON 1992 herausgegebenen Buchs „Ende der biologischen Vielfalt?“. Wie eigentlich ist diese riesige Vielfalt auf der Erde entstanden? Wie hat sich die Biosphäre entwickelt? Eine Erklärung liefert die auf CHARLES DARWIN (1809–1882) zurückgehende Evolutionstheorie, die unter Einbeziehung von genetischen Grundlagen zur sogenannten synthetischen Theorie weiterentwickelt wurde. Danach kann man mindestens fünf Prozesse unterscheiden, die für die Veränderung der Arten und damit für die Evolution verantwortlich sind:
| 1. | Mutation und Rekombination Als Mutation bezeichnet man spontan oder aufgrund bestimmter physikalischer oder chemischer Einwirkung auftretende Veränderungen im Genom. Die Wahrscheinlichkeit für solche Veränderungen ist zwar insgesamt relativ niedrig (da es zahlreiche Reparationsprozesse gibt, die für eine Stabilität der Nucleinsäuren sorgen), da Genome jedoch aus einer sehr großen Zahl von Genen bestehen, ist der Gesamtanteil der Gameten mit mutierten Genotypen hoch. Die genetische Vielfalt durch Mutationen darf nicht unterschätzt werden. Allerdings ist es so, dass die meisten Mutanten eher ungünstig sind und nur relativ wenige günstige Mutationen vorkommen, die dafür sorgen, dass Merkmale entstehen, die ihren Trägern Vorteile bringen. Mutationsraten wirken auf zweierlei Weise auf die Evolution: Sie schaffen einmal neues genetisches Material, das dann den Einwirkungen anderer Evolutionsfaktoren unterliegt und sie verändern zum anderen die Häufigkeit bestimmter Allele im Genpool der Population. |
| Die Wirksamkeit günstiger Mutationen wird erst durch die Rekombination genetischen Materials, also durch sexuelle Vorgänge im weitesten Sinne, wirklich wirkungsvoll in einer Population umgesetzt. Gerade die jüngsten Erkenntnisse der Genetik sprechen dafür, dass die meisten Gene unseres Genoms schon sehr alt sind und dass Vielfalt und Neuerung vor allem durch neue Kombinationen, Verdoppelungen, Umstellungen usw. zustande kommen. |
| 2. | Anpassungsselektion Die Anpassungsselektion ist der Evolutionsfaktor, der von DARWIN als der Motor für die Entstehung der Arten angesehen wurde. Anpassungsselektion heißt, dass bestimmte Individuen einer Population aufgrund ihrer besseren Tauglichkeit mehr Nachkommen haben als andere. Diese unterschiedliche Tauglichkeit oder Eignung der Phänotypen kann sich z. B. auswirken beim Nahrungserwerb, bei der Flucht vor Räubern, bei der Resistenz gegen Parasiten und Krankheitserregern, bei der Resistenz gegen verschiedene Umweltfaktoren, beim Wettbewerb um einen Geschlechtspartner und beim Brutpflegeverhalten. Anpassungsselektion sorgt dafür, dass eine Art sich an ihre Umwelt anpasst oder – anders ausgedrückt – dass sie in ihre Nische passt. Ändert sich die Umwelt, sorgt Anpassungsselektion in einem gewissen Umfang für eine Anpassung an die neuen Verhältnisse. Ist die Änderung zu drastisch, wird dies allerdings normalerweise nicht möglich und die Art wird verdrängt oder stirbt aus. Bei lange anhaltenden gleichen Umweltbedingungen können sich Arten in derselben Form über viele Jahrmillionen erhalten (stabilisierende Evolution). Solche Arten können so alt werden und von verwandten Arten so weit entfernt sein, weil diese alle ausgestorben sind, dass man von lebenden Fossilien spricht. Ein Beispiel dafür wären etwa der Ginkgo-Baum oder die Brückenechse, die nur noch auf einigen kleinen Inseln vor Nord-Neuseeland lebt. |
3. | Zufallsselektion Auch der Zufall – von Evolutionsbiologen auch Gendrift genannt – dürfte bei der Evolution eine entscheidende Rolle spielen. Wie groß der Anteil der Anpassungsselektion im Verhältnis zur Zufallsselektion ist, hängt sicher von den besonderen Bedingungen ab und ist von Fall zu Fall unterschiedlich zu beurteilen. Bis heute ist dies ein Streitpunkt der Evolutionsforscher. Zufälle dürften auf alle Fälle bei kleinen Populationen eine größere Rolle spielen als bei großen, bei der Neubesiedelung von bisher nicht besiedelten Arealen und bei katastrophenbedingten Veränderungen. |
4. | Migration, Genfluss |
| 5. | Isolation Während alle diese genannten Evolutionsfaktoren zunächst nur zu einer Veränderung innerhalb einer Population führen, ist die Isolation, die Verinselung, der Faktor, der zur Auftrennung der Arten und damit letzten Endes zur biologischen Vielfalt führt. |
Evolutionäre Prozesse können auf Inseln wichtiger sein als Einwanderung und Auswanderung. Im Laufe der Erdgeschichte kann die Inselbildung ganz entscheidend zur biologischen Vielfalt beigetragen haben. Denn auch wenn Inseln nach langer Zeit wieder zu Festländern werden, so sind die auf ihnen durch Isolation entstandenen Arten in der Regel so stabil, dass es nicht mehr zu einer Verschmelzung mit den Stammarten kommen kann.
-
Evolutionsfaktoren


Ein besonders gutes und gut untersuchtes Beispiel für „Inselevolution“ sind die Fruchtfliegen des Hawaii-Archipels. Geologen haben das Alter dieser Vulkaninseln bestimmt. Die älteste mit 5,1 Mio. Jahren ist Kauai, die jüngste mit ca. 0,4 Mio. Jahren die große Insel Hawaii mit noch mehreren aktiven Vulkanen. Auf diesen Inseln leben heute etwa 500 (!!) endemische Fruchtfliegenarten, die alle zur Gattung Drosophila gehören. Sie stammen alle von einem Vorfahr ab, der vor über 5 Mio. Jahren nach Kauai gelangte. In der Darstellung wird die Entstehungsgeschichte weniger Arten herausgegriffen. Die durchgezogenen Linien deuten die Migration an, die gestrichelten Linien die Anpassungsselektion auf der neuen Insel.
Wie kann man sich nun die Aufspaltung einer Stammart in viele Arten vorstellen?
Dies soll an einem einfachen Modell aus drei benachbarten Inseln erläutert werden:
| (1) | Eine Insel wird durch eine kleine Kolonie der Art A besiedelt. Der Genpool dieser Teilpopulation ist vom Zufall bestimmt („Zufallsselektion“). |
| (2) | Vom Genpool der Ausgangspopulation isoliert entwickelt sich diese Inselpopulation in Anpassung an ihre neue Umwelt zu Art B. |
| (3) | Durch Stürme oder andere Umweltereignisse gelangt Art B auf eine weitere Insel. |
| (4) | Auf dieser zweiten Insel entwickelt sich B zu C. |
| (5) | Individuen von C besiedeln erneut die erste Insel, können sich jedoch aufgrund genetischer Barrieren mit B nicht mehr vermischen. |
| (6) | C besiedelt auch die dritte Insel. |
| (7) | C bildet auf der dritten Insel Art D. |
| (8) | Art D wird auf die beiden Inseln ihrer Vorfahren vertrieben. |
| (9) | Auf der ersten Insel bildet D die neue Art E. |
| (10) | Usw. usw. |
Durch dieses Wechselspiel kleiner Populationen können isolierte Habitate zu einer starken Artaufspaltung führen, die von Evolutionsbiologen „adaptive Radiation” genannt wird.
Wie schon angedeutet, kann fehlende Konkurrenz auch zum Überdauern sehr altertümlicher Lebensformen führen. So sind Inseln nicht selten zu letzten Refugien von sehr ursprünglichen, heute isoliert stehenden Arten geworden. Viele solcher „lebenden Fossilien“ sind vom Menschen in historischer Zeit ausgerottet worden, wie etwa die Dronten auf Mauritius und La Réunion (ca. 1800) und die Moas auf Neuseeland (ca.1650).
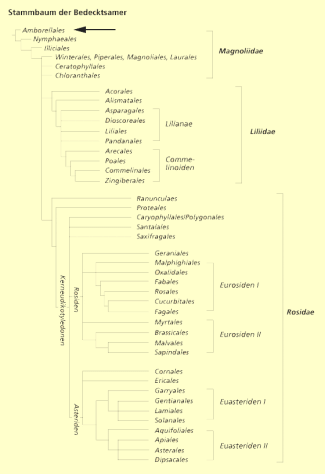
Ein solcher noch überlebender Reliktendemit der Insel Neukaledonien östlich von Australien, die bedecktsamige Pflanze Amborella trichopoda, hat in den letzten Jahren eine gewisse Berühmtheit erlangt. Aufgrund von DNA-Analysen hat man einen Stammbaum der Bedecktsamer entwickelt. Bei diesem Stammbaum steht die erwähnte Art ganz isoliert an der Basis des Systems. Sie ist damit die nächste noch lebende Verwandte der Ursprungsart, die vielleicht vor 150 Millionen Jahren lebte und aus der sich die ganze Vielfalt der „Blütenpflanzen“ von den Wasserlinsen bis zu den Eukalyptusbäumen, rund ¼ Millionen beschriebener Arten, entwickelte. 
Trennung und Verbindung – zur Organisation der Biosphäre
Verinselung ist eine wesentliche Voraussetzung für Biodiversität, allerdings nur, wenn die Isolation nicht vollständig ist, wenn es ein Wechselspiel zwischen Auftrennung und Verbindung geben kann.
Ein solches Wechselspiel hat im größten Maßstab und in erdgeschichtlichen Zeiträumen stattgefunden. Im Laufe der Erdgeschichte hat sich die Verteilung der Festländer und der Ozeane ständig verändert. Als Ursache dieser „Kontinentaldrift“ wurden zyklische Vorgänge in den äußeren Schichten unseres Planeten erkannt, die man mit dem Schlagwort „Plattentektonik“ kennzeichnet. In dieser Zeit sind die Dinosaurier aufgeblüht und wieder untergegangen, die Vögel und die Säugetiere haben sich entwickelt, ebenso die „Blütenpflanzen“. Die heutige Verteilung der Arten und die Biodiversität, so wie sie sich heute darstellt, wurde ganz entscheidend durch diese plattentektonischen Vorgänge bestimmt, die durch Trennung und Verbindung von Festländern und Meeresräumen gekennzeichnet sind.
-
Stammbaum der Bedecktsamer mit Amborella trichopoda als erste Pflanze.

Aber auch im kleinen und kleinsten Maßstab spielen solche Vorgänge der Isolation und der Verbindung eine wichtige Rolle. Von dem Zusammenspiel der Ökosysteme war schon die Rede, auch von der Wechselwirkung von Populationen einer Art. Innerhalb der Populationen sind es die Individuen, die voneinander klar getrennt und genetisch und morphologisch und physiologisch einzigartig doch zur Eigenart und zum Genpool der Gesamtpopulation beitragen. Und die Individuen, die Einzelorganismen? Auch sie sind zusammengesetzt aus mehr oder weniger autonomen Teilen. Wie könnte man Organe transplantieren, wenn es diese Autonomie nicht gäbe?
Selbst die kleinste Einheit des Lebens, die Zelle, stellt sich im elektronenmikroskopischen Bild als ein Kosmos aus vielen Einzelteilen, als eine Landschaft mit Inseln und Seen, man könnte auch sagen als ein dreidimensionales Labyrinth dar. Biologen sprechen von „Kompartimentierung“. Jedes Kompartiment ist eigentlich eine Insel, aber eine Insel, zu der es Fährverbindungen gibt.
Die zwanzig größten Inseln der Erde
| Die größten Inseln der Erde (in km²) | |||
| Grönland | 2 130 800 | Celebes | 189 216 |
| Neuguinea | 771 900 | Neuseeland, Südinsel | 151 757 |
| Borneo | 754 770 | Java | 118 000 |
| Madagaskar | 587 041 | Neuseeland, Nordinsel | 114 597 |
| Baffin Island | 507 541 | Neufundland | 108 860 |
| Sumatra | 425 000 | Kuba | 105 007 |
| Honshu | 231 090 | Luzon | 104 688 |
| Großbritannien | 229 957 | Island | 103 000 |
| Victoria Island | 217 290 | Mindanao | 94 630 |
| Ellesmere Island | 196 236 | Irland | 84 403 |
Suche nach passenden Schlagwörtern
- Naturschutz
- Inselbesiedelung
- Biogeographie
- Sukzession
- Mutation
- Stammbaum
- Isolation
- Theorie der Habitatvielfalt
- Evolutionstheorie
- Relikte
- Biogeografie
- Fruchtfliegen
- Klimaxzustand
- Adaptive Radiation
- Artenspektrum
- Lebensraum
- Fossilien
- Genfluss
- Barriere
- Ökologie
- Buntbarsche
- Mosaik-Zyklus-Theorie
- Reisebericht
- Biodiversität
- Auswanderungsrate
- Ökologische Nische
- Stammesgeschichte
- Anpassungsselektion
- Paläontologie
- Einnischung
- Gleichgewichtstheorie
- Gendrift
- Inselgeographie
- Endemiten
- Einwanderungsraten
- biologische Vielfalt
- Artenzahl
- Zyklen

