Beeinflussung der Fotosyntheseleistung
Die Fotosynthese ist vorwiegend von Licht, Kohlenstoffdioxidkonzentration und Temperatur abhängig, wobei generell der Faktor die Fotosyntheseleistung begrenzt, der im Minimum vorliegt (Gesetz des Minimums von LIEBIG). Die Abhängigkeit der Fotosyntheseleistung von Licht und Kohlenstoffdioxid lässt sich grafisch als Sättigungskurve darstellen. Der Licht- bzw. Kohlenstoffdioxidkompensationspunkt ist für die Pflanzen wichtig, da bei Überschreitung des Kompensationspunkts organische Stoffe innerhalb der Pflanze zur Speicherung von chemischer Energie gebildet werden können. Die Temperaturabhängigkeit der Fotosyntheseleistung wird in Form einer Optimumskurve beschrieben. Die Abhängigkeit der Fotosyntheseleistung von den Faktoren unterscheidet sich zum einen zwischen C3-Pflanzen und Fotosynthesespezialisten (C4-Pflanzen, CAM-Pflanzen) und ebenso bei Licht- und Schattenpflanzen bzw. Licht- und Schattenblättern.
Fotosynthesefaktoren
Die Fotosynthese wird wie alle Lebensprozesse von verschiedenen Bedingungen beeinflusst:
- vom Entwicklungszustand der Pflanze,
- vom Öffnungszustand der Stomata,
- vom Chlorophyllgehalt,
- von der Wasser- und Mineralsalzversorgung,
- von der Qualität und Intensität des Lichts,
- von der Temperatur sowie
- von der Kohlenstoffdioxidversorgung.
Alle diese Faktoren stehen z. T. in einem engen Zusammenhang und bedingen einander. Unter natürlichen Bedingungen liegt die Mehrzahl der Faktoren selten im optimalen Einflussbereich für die Pflanzen und begrenzt dadurch die Fotosyntheseleistung. Die Fotosyntheseintensität wird nach dem Gesetz des Minimums von JUSTUS LIEBIG (1803-1873) durch denjenigen Faktor begrenzt, der am wenigsten vorhanden ist. So können Pflanzen z. B. optimale Licht- und Temperaturverhältnisse für die Fotosyntheseintensität nur bedingt ausnutzen, wenn beispielsweise die Kohlenstoffdioxidversorgung unzureichend ist.
Der Begriff Fotosyntheseintensität wird durch die Stoffproduktion pro Zeiteinheit definiert. Als Messgrößen werden Sauerstoffabgabe, Kohlenstoffdioxidaufnahme oder die Herstellung organischer Stoffe verwendet.
Neben der Fotosynthese laufen gleichzeitig dissimilatorische Prozesse in der Pflanze ab, die Zwischenprodukte der Fotosynthese nutzen. Deshalb wird zwischen Brutto- und Nettofotosynthese unterschieden. Die Beziehung kann folgendermaßen zusammengefasst werden:
Nettofotosynthese = Bruttofotosynthese minus Dissimilation.
Man nimmt heute an, dass ein Quadratmeter grüner Blattfläche unter günstigen Bedingungen 0,5-1,5 g Glucose pro Stunde herstellen kann. Dabei wird so viel Kohlenstoffdioxid verbraucht, wie in drei Kubikmetern Luft vorhanden ist.
Chlorophyllgehalt
Der Chlorophyllgehalt einer Pflanze ist unter natürlichen Bedingungen kein begrenzender Faktor. So können selbst Pflanzen mit geringem Chlorophyllgehalt ausreichend Fotosynthese betreiben. Der Chlorophyllgehalt kann aber bei Pflanzen in Anpassung an unterschiedliche Lichtintensitäten variieren. So weisen Schattenblätter im Vergleich zu Sonnenblättern einen höheren Chlorophyllgehalt pro Einheit Blattfläche auf.
Licht
Die Fotosynthese ist primär von der Lichtenergie abhängig, da durch die lichtabhängigen Reaktionen die Voraussetzungen (ATP, ) für die im CALVIN-Zyklus ablaufenden Prozesse geschaffen werden.
Der Kurvenverlauf für die Abhängigkeit der Fotosyntheseleistung von der Lichtenergie beginnt im negativen Bereich, da hier vorerst der Sauerstoffverbrauch durch die Zellatmung größer ist als die Sauerstoffproduktion durch die Fotosynthese. Solange keine anderen Umweltfaktoren (z. B. Kohlenstoffdioxid, Temperatur) begrenzend wirken, wächst zunächst die Fotosyntheseintensität mit Erhöhung der Lichtintensität proportional. Am Lichtkompensationspunkt, wo die Sauerstoffabgabe (bzw. Kohlenstoffdioxidaufnahme) durch die Fotosynthese genauso so groß ist wie der Sauerstoffverbrauch (bzw. Kohlenstoffdioxidproduktion), schneidet die Kurve die x-Achse und die Nettofotosynthese hat den Wert null.
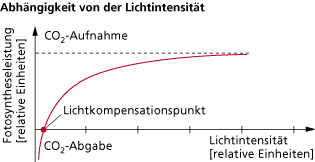
Nur Pflanzen, deren Produktion von Sauerstoff bzw. Verbrauch an Kohlenstoffdioxid über dem Lichtkompensationspunkt liegt (höhere Fotosyntheseleistung als Atmungsintensität), können organische Stoffe speichern. Mit zunehmender Strahlungsintensität wirken die anderen Faktoren nach dem Gesetz des Minimums immer stärker limitierend auf die Fotosyntheseleistung. Dadurch steigt die Kurve der Fotosyntheseleistung immer weniger und zwar bis zu dem Punkt, an dem sich trotz steigender Lichtintensität die Fotosyntheseleistung nicht mehr erhöht (Lichtsättigung).
Der Kurvenverlauf der Fotosyntheseintensität in Abhängigkeit von der Lichtintensität wird dementsprechend als Sättigungskurve bezeichnet. Falls die Lichteinwirkung trotzdem weiter ansteigt, kann dies zu einer Schädigung der Fotosysteme (unter Beteiligung von Sauerstoffradikalen) führen. Unter natürlichen Bedingungen tritt dies z. B. bei sehr geringen Temperaturen oder bei Schattenpflanzen, wenn sie plötzlich sehr starkem Lichteinfluss ausgesetzt werden, auf. In begrenztem Umfang können bestimmte Fotosynthesepigmente (Xanthophylle) überschüssige, nicht verwertbare Lichtenergie zum Schutz vor entstehenden Sauerstoffradikalen auffangen.
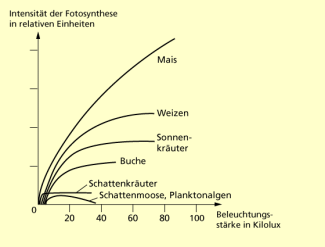
Pflanzenarten zeigen unterschiedliche Anpassungen an die Lichtintensität. So unterscheiden sich Fotosynthesespezialisten, wie der Mais als C4-Pflanze, von den C3-Pflanzen. C4-Pflanzen erreichen selbst bei hoher Lichtintensität keine absolute Lichtsättigung. Deshalb sind C4-Pflanzen den C3-Pflanzen bei voller Sonneneinstrahlung in Bezug auf die Strahlungsverwertung überlegen. Bei C3-Pflanzen unterscheidet man an Starklicht (Sonne) und Schwachlicht (Schatten) angepasste Pflanzen oder Organe. Die Möglichkeiten sind genetisch festgelegt oder variieren infolge von Modifikationen.
Beim Anbau von Pflanzen sollte man u. a. auf die Pflanzabstände achten oder durch zusätzliche Beleuchtung für eine ausreichende Lichtzufuhr sorgen.
Kohlenstoffdioxid
Der Kurvenverlauf für die Abhängigkeit der Fotosyntheseleistung von der Kohlenstoffdioxidkonzentration entspricht ebenfalls einer Sättigungskurve. Die Fotosyntheseleistung steigt bei den meisten Pflanzen proportional mit der Erhöhung des Kohlenstoffdioxidanteils bis auf ca. 0,1 Vol.-%. Danach folgt bei vielen Pflanzen trotz steigender Kohlenstoffdioxidkonzentration keine Zunahme der Fotosyntheseleistung (Kohlenstoffdioxidsättigung) mehr, wobei ein zu hoher Kohlenstoffdioxidanteil auch zu Schädigungen führen kann. Der Kohlenstoffdioxidkompensationspunkt liegt bei C3-Pflanzen zwischen 0,005 und 0,01 Vol.-% Kohlenstoffdioxid in der Atmosphäre.
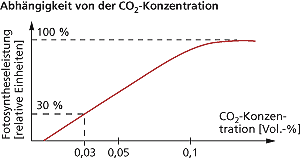
Unter natürlichen Bedingungen ist bei C3-Pflanzen der Kohlenstoffdioxidanteil in der Atmosphäre (0,03 Vol.-%) der begrenzende Faktor für die Fotosyntheseleistung. Die Fotosyntheseintensität kann bei diesen Pflanzen durch Kohlenstoffdioxidanreicherung in Gewächshäusern (Kohlenstoffdioxiddüngung) bis zum Dreifachen gesteigert werden. Ebenso weisen schlecht gelüftete Räume (Kohlenstoffdioxidgehalt ca. doppelt so hoch wie im Freien) oder die Stallmistdüngung diesen Effekt der Erhöhung auf. Letzteres bewirkt zum einen die erhöhte Zufuhr von Mineralstoffen und zum anderen ist die vermehrte Atmungstätigkeit der Mikroorganismen für einen Anstieg der Kohlenstoffdioxidkonzentration verantwortlich, welches sich generell in bodennahen Schichten feststellen lässt. So geben Wald- und Wiesenböden ca. 50 bis 500 ml Kohlenstoffdioxid pro Stunde und Quadratmeter ab. Pflanzen in der Krautschicht nutzen fast ausschließlich diesen erhöhten Kohlenstoffdioxidanteil. Außerdem hemmt der erhöhte Kohlenstoffdioxidanteil der Luft die Fotorespiration und erhöht somit die Nettofotosynthese.
Fotosynthesespezialisten (C4-Pflanzen, CAM-Pflanzen) erhöhen in Anpassung an sonnige trockene Standorte unter Energieverbrauch die Konzentration an Kohlenstoffdioxid, indem sie zunächst einen anderen Akzeptor (PEP) zur Kohlenstoffdioxid-Fixierung verwenden. So können sie auch bei sehr niedrigen Kohlenstoffdioxidkonzentrationen noch eine hohe Fotosyntheseleistung erzielen.
Die Kohlenstoffdioxidversorgung ist für Pflanzen im Wasser weniger problematisch. Kohlenstoffdioxid löst sich bei einer Wassertemperatur von 15 °C etwa im gleichen Prozentsatz wie es auch in der Luft vorliegt. Zusätzlich wird der langsamere Transport von Kohlenstoffdioxid durch die Wasserbewegung ausgeglichen. Untergetauchte Wasserpflanzen nehmen durch anatomische Besonderheiten (keine Cuticula und Spaltöffnungen) über die gesamte Blattoberfläche entweder nur Kohlenstoffdioxid oder zusätzlich noch Calciumhydrogencarbonat auf.
Temperatur
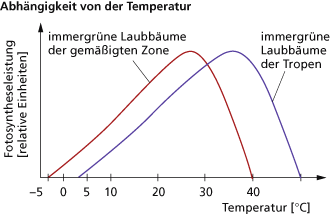
Wie bei allen enzymatisch gesteuerten Reaktionen ist auch die Fotosyntheseleistung von der Temperatur abhängig und wird in Form einer Optimumskurve beschrieben.
-
Abhängigkeit der Fotosyntheseleistung verschiedener Pflanzen von der Beleuchtungsstärke

So steigt die Fotosyntheseleistung bei ausreichender Versorgung mit den anderen Bedingungen entsprechend der RGT-Regel bis zum Optimum (RGT-Regel: Erhöhung der Temperatur um 10 K bewirkt eine Leistungssteigerung auf ca. das Doppelte). Nach Erreichen des Optimums wirken andere Faktoren immer stärker limitierend auf die Fotosyntheseleistung, so u. a. Licht- und Kohlenstoffdioxidversorgung, Erhöhung der Atmungsintensität oder Denaturierung der beteiligten Enzyme. Gleichzeitig haben höhere Temperaturen Einfluss auf den Wassergehalt innerhalb der Pflanze. Durch die erhöhte Transpiration sinkt der Wassergehalt in der Pflanze, sodass sich bei höheren Temperaturen die Spaltöffnungen schließen und so die Kohlenstoffdioxidzufuhr stark behindern. Die Werte der Optimumskurve können innerhalb der Pflanzenart und zwischen den Arten sehr unterschiedlich sein und können durch Temperaturanpassungen verschoben werden:
| Minimum | • • • | höhere Pflanzen gemäßigter und kalter Gebiete in Abhängigkeit der Zellsaftkonzentration: ca. 0 °C immergrüne Nadelhölzer, Wintergetreide, Spinat, Feldsalat: -2 bis -3 °C Flechten: -25 °C Tropenpflanzen: +5 bis +7 °C |
| Optimum | • • | C3-Pflanzen:
C4-Pflanzen: 30-40 °C |
| Maximum | • • | C4-Pflanzen: bis 60 °C Cyanobakterien: >70 °C |
Durch die Verwendung einer Heizung oder geeigneter Kühlmöglichkeiten (z. B. Ventilatoren, Fenster öffnen) können optimale Temperaturbereiche für die Fotosynthese erreicht werden.
Wasser
Wasser ist neben Kohlenstoffdioxid ein direkter Ausgangsstoff für die Fotosynthese. Die benötigte Menge an Wasser ist jedoch so gering, dass sie sich nicht direkt auf den Stoff- und Energiewechselprozess auswirkt. Vielmehr werden indirekt durch den Wassermangel die enzymatischen Prozesse gehemmt und durch den Verschluss der Spaltöffnungen wird die Kohlenstoffdioxidaufnahme entscheidend beeinträchtigt. Die Pflanzen greifen zunächst auf intern entstandenes Kohlenstoffdioxid der Zellatmung zurück, bevor in weiteren Stufen die Kohlenstoffdioxidzufuhr ganz gedrosselt wird und dann Schäden an Fotosyntheseapparat und Membranen auftreten.
Der Wasserbedarf ist je nach Pflanzenart verschieden und kann sich innerhalb bestimmter Grenzen in Anpassung an die Umweltfaktoren verschieben. C3-Pflanzen unterscheiden sich dabei von den Fotosynthesespezialisten, wobei zwischen C4- und CAM-Pflanzen ebenfalls Unterschiede auftreten.
Mineralstoffe
Die Mineralstoffe, die über die Wurzel in die Pflanze aufgenommen werden, sind u. a. für den Ablauf aller biochemischen Prozesse und auch zum Aufbau von Zellstrukturen notwendig. Chloroplasten benötigen Mineralstoffe (Mg, Fe, Cu, Mn) für den Bau des Fotosyntheseapparats. Diese sind außerdem Bestandteil der Pigmente und Redoxsysteme (Elektronentransportkette). Die Primärprodukte der lichtunabhängigen Reaktion werden mithilfe von Mineralstoffen zu anderen organischen Stoffen (Eiweißen) umgewandelt und dabei u. a. auch zur Speicherung von Energie verwendet.
Regelmäßige Düngung sichert eine ausreichende Mineralstoffversorgung für die Pflanzen.

