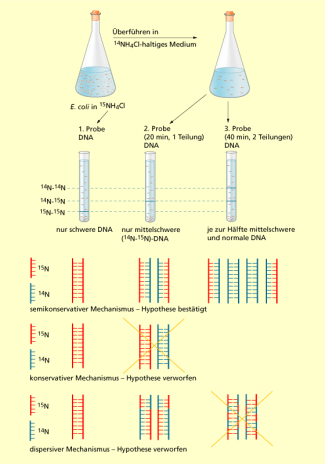
Das MESELSON-STAHL-Experiment
Bei der Vererbung wird das genetische Material der Eltern auf die nachfolgende Generation weitergegeben. Die genetische Information liegt in der DNA in Basenpaaren aus Nucleotiden verschlüsselt vor. Die Synthese der neuen DNA erfolgt durch Auftrennung und identisches Kopieren des Elternstrangs. Jeder Einzelstrang ergänzt seinen jeweils komplementären Strang. Lange war man sich des genauen Mechanismus der DNA-Verdoppelung (Replikation) nicht sicher. Im Jahr 1958 bewiesen MATTHEW MESELSON (*1930) und FRANKLIN WILLIAM STAHL (*1929) den semikonservativen Replikationsmechanismus der DNA, den zuvor WATSON und CRICK, die Erforscher der DNA-Doppel-Helix-Struktur, schon vermuteten. Von drei zur damaligen Zeit möglichen Hypothesen konnten sie in ihrem klassischen Experiment eine bestätigen.
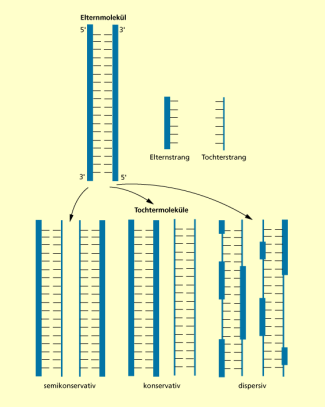
JAMES D. WATSON (*1928) und FRANCIS H. C. CRICK (1916-2004) legten schon 1953 in ihrem Artikel zur DNA-Struktur nahe, dass die komplementären Basenpaarungen wichtig für die Verdoppelung der DNA sind. Sie bezogen sich aber nur darauf, dass die Nucleotide als Matrize/Vorlage zur Replikation dienen können. Über das Grundschema des Replikationsablaufs waren sich die Genetiker Anfang der 1950er-Jahre jedoch noch im Unklaren. Drei unterschiedliche Möglichkeiten waren denkbar:
- Bei der konservativen Replikation besteht das eine Tochtermolekül aus den beiden Elternsträngen und das andere besteht aus zwei neuen Strängen.
- Bei der semikonservativen Replikation besteht jedes Tochtermolekül aus je einem ursprünglichen und einem neuen Strang.
- Bei der dispersiven Replikation bestehen beide Tochtermoleküle aus einem Gemisch von alten und neuen Strängen.
-
Möglichkeiten der Replikation

Der semikonservative Mechanismus war für WATSON und CRICK zwar naheliegend, musste aber durch ein Experiment bewiesen werden. Das Schlüsselexperiment dazu führten MATTHEW MESELSON und FRANKLIN STAHL 1958 am California Institute of Technology durch.
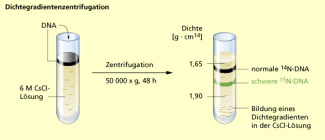
Für ihr Experiment nutzen sie die Masseunterschiede der Stickstoffisotope und . Der häufig vorkommende Stickstoff besitzt eine relative Atommasse von 14,008. Das seltenere Isotop ist um die Masse eines Neutrons (1,0087) schwerer. Da beide Isotope sich chemisch so gut wie gar nicht unterscheiden und nicht radioaktiv ist, kann das schwerere Isotop ohne Weiteres in Form von Nährstoffen dem Stoffwechsel von Bakterien zugeführt werden. Nährstoffe werden von den Bakterien verstoffwechselt und wird u. a. in die Strukturen der DNA eingebaut. Durch Dichtegradientenzentrifugation können die schwereren Stoffwechselprodukte aufgrund ihrer größeren Schwebedichte nachgewiesen werden.
-
Dichtegradientenzentrifugation

MESELSON und STAHL ließen eine Escherichia coli-Kultur zunächst in Anwesenheit von „schwerem“ Ammoniumchlorid wachsen. E. coli baute den in seine DNA ein. Nach der ersten Wachstumsphase wurden die Bakterien in eine Kultur mit „normalem“ Stickstoff umgesetzt. Im neuen Medium wuchsen die Bakterien eine bzw. zwei Replikationsrunden, unter optimalen Bedingungen ungefähr 20 bzw. 40 Minuten. Die DNA-Proben der drei einzelnen E. coli-Kulturen wurden zentrifugiert:
| 1. Probe: | Die Probe, vor dem Umsetzen auf , zeigte eine schwere . Beide Stränge der Doppelhelix enthielten . |
| 2. Probe: | Die Bande der ersten Replikationsrunde zeigte eine mittelschwere DNA zwischen der und der (Blindprobe) an, was einer gemischten entspricht. |
| 3. Probe: | In der Kultur nach der zweiten Replikationsrunde konnten zwei schmalere Banden nachgewiesen werden. Die eine Bande entsprach der , die andere stimmte mit der überein. |
Die Beobachtungen bewiesen eindeutig, dass ein semikonservativer Replikationsmechanismus vorlag. Die DNA der 2. Probe bestand aus je einem alten und einem neuen nach der ersten Replikation. Wenn bei der nachfolgenden zweiten Replikation der als Vorlage diente, entstanden wieder gemischte Moleküle. War der neue die Vorlage, entstanden im wieder komplette Beide Banden waren schmaler, da sie jeweils nur die Hälfte der DNA beinhalteten.
Schon nach der zweiten Probe kann ein konservativer Replikationsverlauf ausgeschlossen werden, hier hätten eine und eine entstehen müssen. Die dispersive Replikation wäre aber noch möglich. Ob die DNA völlig gemischt beide Stickstoffisotope beinhaltet (dispersiv) oder die Isotope auf die Einzelstränge verteilt (semikonservativ) konnte hier nicht unterschieden werden. Durch die dritte Probe konnte der dispersive Mechanismus auch ausgeschlossen werden. Es hätte wieder nur eine gemischte Bande entstehen dürfen.
Mit diesem Beispiel klassischer wissenschaftlicher Arbeit bewiesen MESELSON und STAHL, welcher Replikationsmechanismus bei der Verdoppelung der DNA vorlag. Der präzise Verlauf der Replikation wurde erst später, beginnend in den 1960er-Jahren, aufgeklärt.
Verlauf der Replikation
Bevor die Elternstränge aufgetrennt werden können, müssen sie unter Energieaufwand (ATP-Spaltung) entspiralisiert werden. Die Entschraubung erfolgt durch einen Bruch in einem der Stränge. Die nun geöffnete DNA wird durch Bindung bestimmter Proteine stabilisiert. Beim Auftrennen der beiden Stränge entsteht die Replikationsgabel. Nun lagern sich – unter Mitwirkung von DNA-Polymerasen – die einzelnen Nucleosidtriphosphate durch Wasserstoffbrücken an ihr komplementäres Gegenüber an und werden unter Abspaltung von Diphosphat und Bildung von Phosphatesterbrücken verknüpft. Die Zuordnung der einzelnen Nucleotide ist durch die Basensequenz (Basenabfolge) des Eltern-Einzelstrangs vorgegeben.
-
MESELSON-STAHL-Experiment