DNA-Schäden und DNA-Reparatur
Durch endogene und exogene Faktoren verursacht treten an DNA-Molekülen immer wieder Schäden bzw. Fehler auf. Ebenso können im Verlauf der Replikation des Erbmaterials Fehler entstehen. Derartige Schäden können u. a. sein: fehlende Basen, veränderte Basen, inkorrekte Basenpaarung, Deletion oder Insertion einzelner oder mehrerer Nucleotide in einem der beiden DNA-Stränge, Pyrimidin-Dimere, Strangbrüche (Einzelstrangbruch) oder die kovalente Quervernetzung der DNA-Stränge. DNA-eigene Reparatursysteme dienen der Schadensbehebung und sichern somit das Überleben einzelner Zellen und Organismen. Bei der Replikation entstehende falsche Basenpaarungen können direkt durch die Fehlerbehebungs-Funktion von DNA-Polymerasen eliminiert werden. Im weiteren Verlauf des Zellzyklus auftretende DNA-Schäden können u. a. durch Krebs verursachende Chemikalien, ultraviolette Strahlung oder freie Radikale hervorgerufen werden. DNA-Reparatur-Prozesse sind im gesamten Organismenreich weitverbreitet und verlaufen in allen Organismen ähnlich, aus technischen Gründen sind sie jedoch an Mikroorganismen, besonders Escherichia coli, am gründlichsten untersucht.
Die Replikationsgenauigkeit der DNA bei den Prokaryota ist relativ hoch. Die gesamte Fehlerrate beträgt lediglich 1 Fehler pro Nucleotide, obwohl Fehlpaarungen bei der Elongation sehr viel häufiger sind. Die hohe Genauigkeit wird erzielt durch:
| Fehlerrate danach 1 pro Nucleotide | |
| Fehlerverminderung um das 200- bis 1 000-Fache | |
| verbleibende Fehlerrate 1 pro |

DNA-Schäden
Durch äußere und innere Ursachen können an den großen DNA-Molekülen mannigfaltige Schäden auftreten. Die Replikation oder Transkription kann beeinträchtigt werden. Häufig bewirken die Schäden Mutationen (bleibende Veränderungen des genetischen Materials).
Die wichtigsten Schäden sind:
|
-
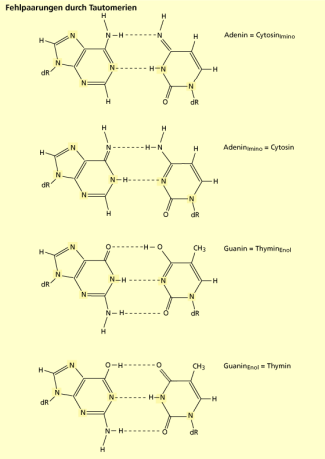
Fehlpaarungen durch Tautomerien
-
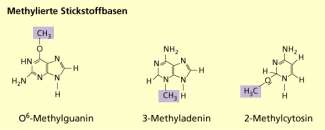
Methylierte Stickstoffbasen
-
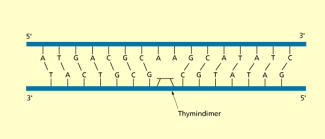
Thymindimer



| |
| |
| |
| |
|
-
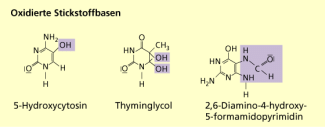
Oxidierte Stickstoffbasen

Bakterielle DNA-Reparatursysteme
Hinsichtlich der vielfältigen möglichen Schäden an DNA gibt es verschiedene Reparatursysteme, mit denen auch Replikationsfehler beseitigt werden. Alle Systeme basieren auf der Redundanz der Information im DNA-Doppelstrang, dem mehrfachen Vorhandensein der Information. Es werden vier grundsätzliche Reparaturmechanismen unterschieden:
- Fehler im Zuge der Replikation werden durch die Fehlpaarungsreparatur behoben. Enzyme kontrollieren während der Replikation die komplementären Basenpaare. Ein erkannter Fehler wird sofort auf dem Tochterstrang korrigiert. Der Elternstrang, sozusagen das Original der Kopie, wird durch modifizierte Adenin-Nucleotide erkannt.
- Durch direkte Reparatur werden beispielsweise Thymin-Dimere beseitigt. DNA-Fotolyasen, Enzyme, die bei Licht aktiviert werden, reparieren den Schaden.
Ebenso direkt können Methylierungen der Nucleotide beseitigt werden. DNA-Reparatur-Methyltransferasen binden in ihrem aktiven Zentrum die überschüssige Methylgruppe. Die DNA-Base ist wieder im Originalzustand. Da die Methylgruppe sich nicht mehr aus dem Enzym lösen kann, wird es unbrauchbar. Die inaktiven Enzyme steigern aber über genregulative Prozesse ihre Neusynthese.
Dieser Mechanismus wird zwar nicht sehr häufig genutzt, ist aber im Falle der genannten Beispiele effizient. - Die Reparatur durch Ausschneiden ( Excision ) ist weitverbreitet. Je nachdem, ob eine einzelne Base oder eine kurze DNA-Sequenz ausgeschnitten werden, unterscheidet man Basen- oder Nucleotid-Excisionsreparatur. Prinzipiell unterscheiden sich beide Mechanismen nicht; die beteiligten Enzyme unterscheiden sich jedoch. Nach dem enzymatischen Erkennen des Fehlers wird er herausgeschnitten und anschließend durch erneute komplementäre Basenpaarung repariert.
- Große Schäden wie Doppelstrangbrüche oder stark beschädigte, verformte Elternstränge können, wenn möglich, durch die Rekombinationsreparatur behoben werden.
Beide Schäden führen zu Störungen der Replikation. Im ersten Fall bricht die Replikation ab, es entstehen DNA-Fragmente, und im zweiten Fall fehlt die Vorlage für den Tochterstrang, sodass die Replikation erst an intakten Sequenzen weiter verläuft.
Zur Reparatur werden homologe DNA-Sequenzen als Vorlage für den intakten Zustand benötigt. Da sich schnell teilende Bakterienzellen meist mit einer neuen Replikationsrunde beginnen, bevor die erste abgeschlossen ist und sich die Zelle geteilt hat, befinden sich mehrere Kopien der DNA in einer Zelle, die als Vorlage dienen können. Durch komplizierte Rekombinationsprozesse (Auffinden homologer Abschnitte, Kombination, Synthese) werden die Schäden repariert. Der Reparaturerfolg hängt jedoch stark von der Schadensgröße ab.
Eukaryotische DNA-Reparatur
Bei Eukaryota erfolgt die Reparatur der DNA mit großer Wahrscheinlichkeit nach den gleichen Prinzipien wie bei den Prokaryota. Allerdings sind sie hier aufgrund des größeren Genoms und seiner komplizierten Struktur wesentlich komplexer.

