Lichtreaktionen – genauer betrachtet
Die Fotosynthese besteht aus zwei übergeordneten Vorgängen, die jede Menge Teilreaktionen enthalten: den Lichtreaktionen (der Foto- bzw. Lichtteil der Fotosynthese) und dem CALVIN-Zyklus (auch Dunkelreaktionen – der Syntheseteil der Fotosynthese).
Innerhalb der Lichtreaktionen werden die stofflichen Voraussetzungen (ATP, NADPH + H+) für die darauffolgenden lichtunabhängigen Reaktionen gebildet. Die Lichtreaktionen finden an den Thylakoidmembranen der Chloroplasten statt, die dafür besondere Strukturen aufweisen: Fotosystem I und II, Elektronentransportkette sowie das Enzym ATP-Synthase.
ROBERT EMERSON (1903-1959) fand durch den nach ihm benannten EMERSON-Effekt experimentell heraus, dass zwei Fotosysteme existieren und damit zwei Lichtreaktionen zusammenwirken.
Fotosysteme absorbieren mithilfe der Fotosynthesepigmente die Lichtenergie. Die absorbierte Lichtenergie wird innerhalb des Fotosystems zu einem Reaktionszentrum weitergeleitet. Dort werden Elektronen auf ein höheres Energieniveau gebracht und stehen für zwei verschiedene Elektronentransporte zur Verfügung.
Beim nichtzyklischen Elektronentransport gelangen die Elektronen von einem Akzeptor des Fotosystems II über eine Elektronentransportkette zum Fotosystem I und daher nicht zu ihrem Ursprungsort zurück. Beim Transport der Elektronen wird mithilfe der Chemiosmose ATP gebildet (nichtzyklische Fotophosphorylierung). Am Fotosystem II findet die Fotolyse des Wassers statt, sodass neben Elektronen und Protonen auch Sauerstoff entsteht. Diese Protonen reagieren später zusammen mit den energiereichen Elektronen des Fotosystems I zum Reduktionsmittel NADPH + H+.
Beim zyklischen Elektronentransport ist nur das Fotosystem I beteiligt. Die durch Lichtabsorption entstehenden energiereichen Elektronen kehren zu ihrem Ursprungsort zurück. Dabei entsteht zusätzlich ATP.
Die Fotosynthese als Form der autotrophen Assimilation umfasst zwei Teilreaktionen: die lichtabhängigen und lichtunabhängigen (CALVIN-Zyklus) Reaktionen.
Die lichtabhängigen Reaktionen laufen an den Thylakoidmembranen des Chloroplasten ab, die besondere Strukturen enthalten: Elektronentransportkette (Plastochinon, Cytochrom-Komplex, Plastocyanin), Fotosystem I und II und das Enzym ATP-Synthase.
Die beiden Fotosysteme bestehen aus einem Reaktionszentrum, dem primären Elektronenakzeptor und dem Antennenkomplex (LHC: L ight H arvesting C omplex). Mehrere Hundert Fotosynthesepigmente (vor allem Chlorophylle, Carotinoide) wirken im Antennenkomplex zusammen, um möglichst viel Licht im sichtbaren Bereich zu absorbieren. Die unterschiedliche Zusammensetzung der Pigmente garantiert ein breites Absorptionsspektrum. Dabei ergänzen sich Chlorophyll a und b, indem Chlorophyll b Lichtenergie der nicht vom Chlorophyll a erfassten Wellenlängen direkt an Chlorophyll a weiterleitet. Carotinoide haben vor allem die Funktion des Lichtschutzes. Sie übertragen keine Energie auf Chlorophyll a, sondern absorbieren und verteilen überschüssige Lichtenergie, die sonst das Chlorophyll schädigen würde (ähnlicher Effekt wie im menschlichen Auge).
Die Pigmente im Antennenkomplex (Antennenpigmente) sind so um das Reaktionszentrum angeordnet, dass die Größe der aufnehmbaren Energiequanten zum Reaktionszentrum hin abnimmt. Einmal absorbierte Lichtquanten gelangen damit früher oder später zum Reaktionszentrum.
Die Reaktionszentren bestehen aus speziellen Chlorophyll a-Molekülen, die nach der Wellenlänge benannt werden, bei der sie optimal Licht absorbieren. Das Reaktionszentrum des Fotosystems I ist das Pigment 700 (P 700), weil es Licht im Bereich von 700 nm Wellenlänge am besten absorbiert. Im Fotosystem II liegt das Pigment 680 (P 680) als Reaktionszentrum vor. Die Reihenfolge der Benennung beider Fotosysteme in Fotosystem I und II erfolgte nach der Reihenfolge ihrer Entdeckung. Das Fotosystem II existiert jedoch nur bei Organismen, die am Ende ihres fotosynthetischen Stoffwechsels Sauerstoff freisetzen. Diesbezüglich unterscheidet sich beispielsweise die Fotosynthese der Bakterien durch einige Besonderheiten (s. Fotosynthese der Bakterien).
ROBERT EMERSON (1903-1959) fand experimentell heraus, dass bei der ersten Phase der Fotosynthese zwei Fotosysteme und damit zwei Lichtreaktionen zusammenwirken. Wenn Pflanzen gleichzeitig mit langwelligem Licht (z. B. 700 nm) und kurzwelligem Licht (z. B. 650 nm) bestrahlt werden, führt das zu einer deutlich größeren Fotosyntheseintensität als wenn man beide Wellenlängen der Pflanze nur einzeln darbietet. Diese Steigerung der Fotosyntheseleistung wird nach dem Entdecker EMERSON-Effekt genannt.
Durch die absorbierte Lichtenergie, die über die Antennenpigmente aufgenommen und bis zum Reaktionszentrum weitergeleitet wurde, werden in den speziellen Chlorophyll a-Molekülen der Reaktionszentren (P 680 und P 700) die Elektronen angeregt, als Folge auf ein höheres Energieniveau gehoben und schließlich von einem primären Akzeptor aufgenommen. (Der primäre Elektronenakzeptor beim Fotosystem II heißt Pheophytin; beim Fotosystem I ist er ein Chlorophyll a.) Der primäre Akzeptor bewirkt, dass die energiereichen Elektronen vom höheren Energieniveau nicht sofort wieder in den energiearmen Grundzustand zurückfallen (sonst entsteht nur Fluoreszenz = Nachleuchten), sondern später über eine Elektronentransportkette weitertransportiert werden können.
Je nach dem Transportweg der Elektronen unterscheidet man zwischen nichtzyklischem und zyklischem Elektronentransport.
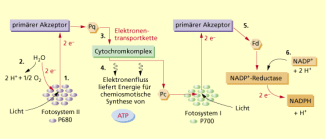
Am nichtzyklischen Elektronentransport sind beide Fotosysteme beteiligt, wobei die Elektronen nicht an ihren Ursprungsort zurückkehren.
Reaktionsschritte:
1. Durch Lichtabsorption gelangen vom Reaktionszentrum (Chlorophyll a-Moleküle – P 680) des Fotosystems II Elektronen auf ein höheres Energieniveau, die vom primären Akzeptor eingefangen werden. Dabei wird pro Chlorophyll a-Molekül nur ein Elektron durch eine Redoxreaktion abgegeben. Das entstandene oxidierte Chlorophyll a-Molekül (P 680) ist als starkes Oxidationsmittel bestrebt, seine Elektronenlücke wieder aufzufüllen.
2. Wasser wird enzymatisch gespalten (Wasser spaltender Komplex). Bei dieser Fotolyse des Wassers (lichtinduzierte Wasserspaltung) entstehen neben Elektronen, die die Elektronenlücke des P 680 im Fotosystem II wieder auffüllen, Protonen und ein Sauerstoffmolekül (). Der bei der Fotosynthese frei werdende Sauerstoff ist also ein Produkt dieses Reaktionsschritts der Fotosynthese.
3. Vom primären Akzeptor des Fotosystems II ausgehend werden die energiereichen Elektronen durch abwechselnde Redoxreaktionen über eine Elektronentransportkette (Plastochinon [Pq] – Cytochromkomplex – Plastocyanin [Pc]) zum Fotosystem I weitertransportiert. Dabei wird mithilfe der Sonnenenergie zu NADPH reduziert. Durch den Wasserstoff wird ein Elektronenpaar auf den Akzeptor übertragen, das zweite geht als Proton in Lösung. Daraus resultiert die richtigere Schreibweise: NADPH + H+. NADPH stellt nun mit der Ansammlung energiereicher Elektronen die erste Speicherquelle der umgewandelten Sonnenenergie dar.
4. Beim Transport der Elektronen wird Lichtenergie in chemische Energie in Form von ATP umgewandelt und gespeichert. Diese Anlagerung von Phosphatgruppen an Adenosindiphosphat (ADP) unter Bildung von ATP mithilfe der Lichtenergie nennt man Fotophosphorylierung und nach dem Weg der Elektronen nichtzyklische Fotophosphorylierung.
5. Parallel zum Fotosystem II werden auch im Reaktionszentrum des Fotosystems I (P 700) durch Lichtabsorption Elektronen angeregt, auf ein höheres Energieniveau gehoben und von einem primären Akzeptor aufgenommen. Die dadurch entstandene Elektronenlücke im oxidierten P 700 wird durch die Elektronen vom Fotosystem II aus der Elektronentransportkette wieder aufgefüllt.
6. Die angeregten Elektronen werden vom primären Akzeptor vom Fotosystem I über Ferredoxin (Fd) auf das Coenzym übertragen, das zusammen mit den Protonen aus der Fotolyse des Wassers zu reagiert.
-
Nichtzyklischer Elektronentransport

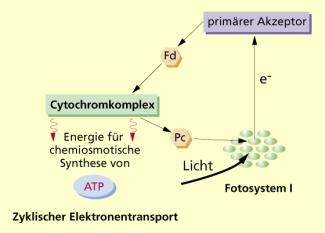
Wenn die Elektronen nach der Lichtabsorption an ihren Ursprungsort wieder zurückkehren, dann wird das als zyklischer Elektronentransport bezeichnet. Dabei ist nur das Fotosystem I beteiligt.
Die durch Lichtabsorption energiereichen Elektronen gelangen über Ferredoxin und Teile der Elektronentransportkette (Cytochrom-Komplex und Plastocyanin) wieder an das Reaktionszentrum P 700 zurück und schließen so die entstandene Elektronenlücke selbst. Dabei findet keine Fotolyse des Wassers und damit weder eine Sauerstoffproduktion noch NADPH + H+-Bildung statt. Als einziges Produkt entsteht Adenosintriphosphat (ATP). Benannt nach der Form des Elektronentransports wird diese Möglichkeit der ATP-Bildung auch als zyklische Fotophosphorylierung bezeichnet.
Durch die Reaktionen des nachfolgend ablaufenden CALVIN-Zyklus ist der ATP-Bedarf höher als durch die nichtzyklische Fotophosphorylierung allein bereitgestellt werden kann. Deshalb steht bei einer genügend hohen -Konzentration diese zusätzliche Möglichkeit der ATP-Bildung zur Verfügung.
Als Ergebnis der lichtabhängigen Reaktionen der Fotosynthese entstehen ATP und NADPH + H+. Dabei stellt ATP die Energiequelle und das Reduktionsmittel für die in der lichtunabhängigen Reaktion (CALVIN-Zyklus) stattfindenden Prozesse dar.
Die Gesamtbilanz der lichtabhängigen Reaktionen lässt sich durch folgende Gleichung zusammenfassen:
Viele Herbizide, die in der Landwirtschaft zur Unkrautbekämpfung eingesetzt werden, greifen an der Elektronentransportkette an. Damit können die lichtabhängigen Reaktionen nicht mehr ausgeführt werden und die Fotosynthese kommt zum Erliegen.
-
Zyklischer Elektronentransport

Suche nach passenden Schlagwörtern
- Video
- Photosynthese
- nichtzyklischen Elektronentransport
- Chloroplasten
- Elektronentransportkette
- Fotosysteme
- EMERSON-Effekt
- Fotolyse des Wassers
- Fotosynthese
- zyklischer Elektronentransport
- Antennenkomplex
- Reaktionszentren
- Fotophosphorylierung
- Thylakoidmembran
- Gesamtbilanz
- Robert Emerson
- Lichtreaktionen

