Fotosynthese
Die Fotosynthese gehört zum aufbauenden Stoffwechsel. Innerhalb ihres Ablaufs werden mithilfe von Lichtenergie aus anorganischen energiearmen Stoffen organische energiereiche Stoffe hergestellt. Chemoautolithotrophie (Chemosynthese) ist eine andere Form des aufbauenden Stoffwechsels. Die Nutzung der Sonnenenergie durch Fotosynthese ermöglichte erst das Leben auf der Erde. Die Chloroplasten sind die Orte der Fotosynthese.
Die Fotosynthese kann man in lichtabhängige und lichtunabhängige Teilreaktionen untergliedern.
Licht, Wasserversorgung, Temperatur und Kohlenstoffdioxidangebot beeinflussen die Fotosyntheseleistung.
| Die Fotosynthese | |
|
|
|
|
|
|
|
|
|
|
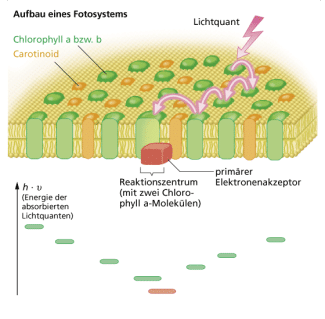
Durch die Fotosynthese wird Sonnenenergie genutzt, um aus Kohlenstoffdioxid und Wasser Kohlenhydrate zu synthetisieren. Ein Teil der Prokaryoten und Protisten sowie die Pflanzen sind auf der Basis dieser Assimilation in der Lage, alle körpereigenen Stoffe aus anorganischen Stoffen aufzubauen. Die Bilanzgleichung der Fotosynthese stellt die Umkehrung der Zellatmung dar:
Die Orte der Fotosynthese in den Zellen der Pflanzen und Protisten sind die Chloroplasten, die auf endosymbiontische Prokaryoten zurückgehen.
Ort der Fotosynthese – Feinbau der Chloroplasten
Chloroplasten sind gegen das Cytoplasma durch eine Doppelmembran abgegrenzt. Die wenig strukturierte Grundsubstanz in ihrem Inneren wird Stroma genannt. Sie enthält Ribosomen, Enzyme und eine ringförmige DNA. Zur Oberflächenvergrößerung ist die innere Membran lamellenartig eingestülpt (Thylakoide). Geldrollenartige Stapel der Thylakoidmembranen werden als Granathylakoide („Körner“), einfache Lamellen im Bereich des Stromas als Stromathylakoide bezeichnet. Die Fotosynthese lässt sich in eine lichtabhängige und eine lichtunabhängige Reaktionsfolge untergliedern. Die Fotosynthesepigmente liegen auf den Granathylakoiden.
Die Thylakoidmembranen mit einer Elektronentransportkette (z. B. Cytochrome), die ATP-Synthase als Enzym und die beiden Fotosysteme ermöglichen die lichtabhängigen Reaktionen. Die lichtunabhängige Reaktion (Calvin-Zyklus) findet im pigmentlosen Stroma statt.
| Chlorophylle | Carotinoide | |
| grüner Blattfarbstoff | gelbe oder rote Blattfarbstoffe | |
| Bau | - 4 Pyrrolringe bilden Porphyrin-Grundgerüst - Mg als Zentralatom - Phytolrest - Unterscheidung durch Reste in ~a, ~b usw. | - kettenförmige Kohlenwasserstoffe mit vielen konjugierten Doppelbindungen |
| Funktion | - Chlorophyll a: Moleküle bilden Reaktionszentrum in Fotosystemen. - Andere Chlorophyllmoleküle sind Hilfspigmente im Antennenkomplex. | - Hilfspigmente im Antennenkomplex - Schutz der Chlorophylle |
Pflanzen nutzen nur die Wellenlängen des sichtbaren Lichts zur Fotosynthese. Die im Chloroplasten vorkommenden Farbstoffe (Pigmente) stellen die Lichtenergie durch Absorption den Pflanzen zur weiteren Verwendung zur Verfügung. Chlorophylle und Carotinoide absorbieren vorwiegend Wellenlängen des blauen und roten Bereichs. So entsteht im mittleren Bereich des sichtbaren Lichts eine „Grünlücke“. Für eine optimale Lichtausbeute bilden die verschiedenen Pigmente einen Fotosystem-Komplex, der sich in den Thylakoidmembranen befindet.
-
Feinbau der Thylakoidmembran
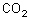

Aufbau der Fotosysteme
Fotosysteme bestehen aus einem Reaktionszentrum und einem Antennenkomplex (LHC = Light Harvesting Complex). Mehrere Hundert Chlorophyll- und Carotinoid-Moleküle bilden einen Antennenkomplex. Dort sind die Pigmentmoleküle so angeordnet, dass die äußeren Pigmente die energiereichsten Photonen absorbieren und nach innen die Größe der aufzunehmenden Energiequanten abnimmt. So landen schließlich alle Energiequanten beim Reaktionszentrum des Fotosystems I oder II, das aus zwei Chlorophyll a-Molekülen besteht. Sie absorbieren die energieärmsten Lichtquanten (680 bzw. 700 nm), wodurch Elektronen angeregt werden. Dadurch erhöht sich die Elektronegativität der beiden Chlorophyllmoleküle und ein Elektron wird an einen primären Akzeptor weitergegeben, bevor es wieder in einen energieärmeren Zustand zurückfallen kann.
Ablauf der Fotosynthese – lichtabhängige Reaktionen
Vom primären Akzeptor werden die Elektronen an mehrere hintereinandergeschaltete Redoxsysteme weitergegeben. Dafür kommen zwei Wege infrage, ein zyklischer und ein nichtzyklischer Elektronentransport. Die Begriffe beziehen sich darauf, ob die vom Reaktionszentrum abgegebenen Elektronen an ihren Ursprungsort zurückkehren oder nicht.
-
Nichtzyklischer Elektronentransport

- Durch Absorption von Lichtenergie wird die Elektronegativität des Chlorophyllmoleküls (Reaktionszentren der Fotosysteme) erhöht, das Redoxpotenzial wird vom positiven in den negativen Bereich verschoben. Dadurch gibt das Chlorophyllmolekül des Reaktionszentrums 2 Elektronen an einen primären Akzeptor ab.
- Das Redoxpotenzial des P 680 ist nun so positiv (+ 810 mV), dass es dem Wasser Elektronen entziehen kann (Fotolyse des Wassers).
- Das vom angeregten P 680 abgegebene Elektronenpaar wird über eine Elektronentransportkette [Plastochinon (Pq) – Cytochromkomplex – Plastocyanin (Pc)] zum Fotosystem I (P 700) weitergeleitet.
- Die Elektronentransportkette liefert die Energie für die ATP-Bildung (nichtzyklische Fotophosphorylierung).
- Das angeregte P 700 gibt 2 Elektronen über Ferredoxin an NADP+ ab, wo es zusammen mit 2 H+ aus der Fotolyse des Wassers zu NADPH + H+ reagiert.
- Bei größerem ATP-Bedarf läuft am Fotosystem I zusätzlich ein zyklischer Elektronentransport ab, der zur ATP-Bildung führt (zyklische Fotophosphorylierung).
Lichtunabhängige Reaktion (Dunkelreaktion, Calvin-Zyklus)
ATP und NADPH + H+ aus den lichtabhängigen Reaktionen sind die Voraussetzungen, dass diese Reaktionsfolge im pigmentlosen Stroma ablaufen kann.
Ablauf der lichtunabhängigen Reaktion (Calvin-Zyklus):
- Fixierung des Kohlenstoffdioxids: Kohlenstoffdioxid wird an Ribulose-1,5-bisphosphat (RuBP) gebunden. Die entstandene instabile Zwischenverbindung zerfällt sofort in 2 Moleküle Glycerinsäure-3-phosphat. Alle Pflanzen, die Glycerinsäure (Verbindung mit 3 C-Atomen im Molekül) als erstes stabiles Zwischenprodukt in der lichtunabhängigen Phase bilden, werden als C3-Pflanzen bezeichnet.
- Reduktion: Jedes Molekül Glycerinsäure-3-phosphat wird zu Glycerinaldehyd-3-phosphat (G3P) reduziert. ATP liefert für diesen energieaufwendigen Prozess die Energie und NADPH + H+ die Elektronen. Aus jeweils zwei Molekülen Glycerinaldehyd-3-phosphat bildet sich unter Abspaltung der Phosphatgruppen Glucose. Daraus kann sich durch Polymerisation Stärke bilden.
- Regeneration des Ribulose-1,5-bisphosphats: Aus jeweils 5 G3P-Molekülen werden 3 Moleküle RuBP regeneriert. Für jeweils 6 CO2-Moleküle, die in den Zyklus eingeschleust werden, entsteht ein Glucose-Molekül. Dafür werden 18 ATP und 12 NADPH + H+ verbraucht.
Wenn man alle Prozesse der lichtabhängigen und lichtunabhängigen Reaktionen zusammenfasst, ergeben sich folgende Gesamtgleichungen:
Beeinflussung der Fotosynthese
Die Fotosyntheseleistung ist vorrangig von der Energie des Sonnenlichts, der Temperatur und dem Kohlenstoffdioxidgehalt der Luft abhängig, wobei jede Art ihre spezifischen Ansprüche an die Umwelt hat.
Dabei stehen die Landpflanzen vor einer prinzipiellen Schwierigkeit: Der geringe -Gehalt der Atmosphäre zwingt sie zur Oberflächenvergrößerung, die Gefahr des Austrocknens durch zu hohe Transpirationsraten wird dadurch erhöht. Teilweise wird dieses Problem dadurch gemildert, dass der Gasaustausch durch die Spaltöffnungen reguliert werden kann.
Beeinflussung der Fotosyntheseleistung
Bei einer Erhöhung der Lichtintensität steigt zunächst die Fotosyntheseleistung an. Am Lichtkompensationspunkt besteht ein Gleichgewicht zwischen -Abgabe und -Aufnahme. Wenn eine weitere Steigerung der Lichtintensität zu keiner weiteren Erhöhung der Fotosyntheseleistung führt, ist die Lichtsättigung erreicht.
Kohlenstoffdioxid ist in der Natur der begrenzende Faktor für die Fotosyntheseleistung. Die Erhöhung der Kohlenstoffdioxidkonzentration bis ca. 0,1 Vol.-% führt zu einer Steigerung der Fotosyntheseleistung. Danach erfolgt trotz steigenden Kohlenstoffdioxidanteils keine Zunahme der Fotosyntheseleistung mehr (Kohlenstoffdioxidsättigung).
-
Abhängigkeit der Fotosyntheseleistung vom Kohlenstoffdioxidgehalt
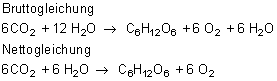

Wie bei allen chemischen Reaktionen steigt die Fotosyntheseleistung bei einer Erhöhung der Temperatur um 10 K auf das Zwei- bis Dreifache (RGT-Regel). Bei mehr als 40 °C sinkt die Fotosyntheseleistung allerdings durch die Denaturierung der an den Reaktionen beteiligten Enzyme. Aufgrund der erhöhten Transpiration schließen sich die Spaltöffnungen bei den Pflanzen, was zur verminderten Kohlenstoffdioxidaufnahme führt.
-
Abhängigkeit der Fotosyntheseleistung von der Temperatur

Teilweise sind die Laubblätter einer Pflanzenart an unterschiedliche Lichtverhältnisse angepasst (Licht- und Schattenblätter, z. B. bei der Rotbuche, Bild 9). Wenn Pflanzen trockener, heißer und sonniger Standorte ihre Spaltöffnungen geschlossen halten, sinkt die -Konzentration in den chlorophyllhaltigen Blattzellen ab, die Sauerstoffproduktion läuft aber noch weiter. Dies kann dazu führen, dass statt an Ribulose-1,5-bisphosphat gebunden wird, was schließlich zur Freisetzung von (Lichtatmung) führt. Dieser Prozess kann durch besondere physiologische Anpassungen verringert werden: C4-Pflanzen (z. B. Mais, Hirse-Arten) binden sehr wirkungsvoll in den Mesophyllzellen, wo als erstes stabiles Produkt ein C4-Körper (z. B. Apfelsäure) entsteht. Von dort erfolgt der Weitertransport der C4-Verbindung über Plasmodesmen in die Gefäßbündelscheidenzellen, wo abgespalten und dem Calvin-Zyklus zur Verfügung gestellt wird. Pflanzen mit tagesperiodischem (diurnalem) Säurezyklus (CAM-Pflanzen) binden das nachts als organische Säure. Aus dieser wird es tagsüber bei geschlossenen Spaltöffnungen wieder freigesetzt.
-
Blattquerschnitt von Licht- und Schattenblättern
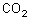

Suche nach passenden Schlagwörtern
- Video
- Chloroplasten
- Fotosysteme
- Stromathylakoide
- C4-Pflanzen
- Antennenkomplex
- Granathylakoide
- Farbstoffe
- Absorption
- Fotosynthesepigmente
- Lichtreaktionen
- Photosynthese
- Elektronentransport
- Elektronentransportkette
- Kohlenstoffdioxidkonzentration
- Carotinoide
- Plasmodesmen
- Dunkelreaktionen
- Fotosyntheseleistung
- Reaktionszentrum
- Temperatur
- Fotosynthese
- CAM-Pflanzen
- Stoffwechsel
- Assimilation
- Chlorophylle
- Stroma
- Granum
- Lichtatmung
- Grana
- Laubblätter
- Lichtintensität
- Pigmente
- Lichtsättigung
- Kohlenstoffdioxidsättigung
- RGT-Regel

