Moose
Moose sind die ursprünglichsten Landpflanzen (Pflanzen im engeren Sinne), die ihren Grünalgenvorfahren noch relativ nahe stehen. Im Gegensatz zu Farnpflanzen und Samenpflanzen haben ihre Vegetationskörper keine Wurzeln. Ihre Blättchen sind den Blättern der übrigen Pflanzen nicht homolog, oft bestehen sie nur aus einer Zellschicht.
Sie können mit dem gesamten Vegetationskörper Wasser aufnehmen. Da den Moospflänzchen schützende Kutikula und Wachsschichten weitgehend fehlen, können sie aber auch durch Verdunstung ziemlich schnell austrocknen. Im Gegensatz zu fast allen anderen Pflanzen können sie diesen Trockenzustand jedoch mehr oder weniger lange in einer Art Scheintod überdauern und nach Befeuchtung wieder aufleben.
Man unterscheidet die Verwandtschaftsgruppen Lebermoose, Hornmoose und Laubmoose.
Moose
Die ersten Landpflanzen
Beim Übergang zum Landleben wurden das lebensnotwendige Wasser und das Kohlenstoffdioxid zu Mangelfaktoren, obwohl man davon ausgehen kann, dass vor einer halben Milliarde Jahre der Kohlenstoffdioxidanteil der Atmosphäre noch wesentlich höher war als heute. Moose und Farne können eine Vorstellung davon vermitteln, welche Verhältnisse zu Beginn der Landpflanzenentwicklung für die Besiedlung der Festländer Voraussetzung und Bedingung waren.
Zu den Einrichtungen, die das Landleben erst ermöglichten, gehören:
- Leitgewebe für Wasser und Assimilatetransport und damit verbunden die Arbeitsteilung zwischen wasseraufnehmenden Organen (bei den Gefäßpflanzen Wurzel mit Wurzelhaaren) und der Assimilation dienenden Organen (bei den Gefäßpflanzen beblätterte Sprosse).
- Festigungsgewebe für die Stabilisierung der Luftsprosse vor allem zur Erhöhung ihrer Biegungsstabilität. Eine besonders wichtige chemische Neuerung in dieser Hinsicht war die Erfindung des Lignins und die damit verbundene Lignifizierung von Zellwänden.
- Schutzeinrichtungen gegen übermäßigen Wasserverlust bzw. Regulationseinrichtungen für die Transpiration und den Gasaustausch, d. h. die Bildung einer geschlossenen Außenhaut (Epidermis) mit einem möglichst dicht schließendem Überzug aus Kutin, einem wachsähnlichen Kohlenwasserstoff, und ihre Durchlässigkeit regulierende Spaltöffnungen.
- Behälter für Keimzellen und Sporen, die gut gegen Austrocknung geschützt sind, mit Schutzhüllen aus unfruchtbaren Zellen sowie besonders austrocknungsresistente Sporenhüllen für die Luftverbreitung der Sporen.
- Embryonenbildung: Vor Austrocknung durch besondere Strukturen geschützte Zygoten und frühere Entwicklungsstadien der Sporophyten.
- Pilzwurzelbildung (Mykorrhiza): Symbiose mit Pilzen, wodurch die Aufnahme von Mineralstoffen und Wasser aus dem Boden erleichtert wurde.
Parallel mit dem Übergang zum Landleben entwickelte sich auch der für (Land)-pflanzen typische Generationswechsel. Die Grünalgenvorfahren der Landpflanzen waren Haplonten, das heißt, sie bestanden aus haploiden Zellen. Die aus der Befruchtung hervorgegangene diploide Zygote teilte sich sofort wieder meiotisch in vier haploide Zellen, aus denen neue haploide Algen entstanden. Dadurch, dass die Zygoten sich nicht sofort wieder durch Meiose (Reduktionsteilung) in haploide Zellen teilen, sondern durch mitotische Teilungen zu einem vielzelligen Gebilde aus diploiden Zellen heranwachsen, entsteht eine zweite Generation. Sie wird Sporophyt genannt, denn von dieser Generation werden als Verbreitungseinheiten durch meiotische Teilungen haploide Sporen gebildet, aus denen sich ohne Befruchtung durch Mitosen direkt eine neue haploide Generation entwickelt. Diese Generation wird Gametophyt genannt, weil sie haploide Gameten produziert.
Im Laufe der Landpflanzenentwicklung haben sich nur die Sporophyten weitgehend den genannten Forderungen angepasst. Die Gametophyten dagegen sind in vieler Hinsicht noch wassergebundene Pflanzen geblieben. Teilweise wird dies durch besondere physiologische Eigenschaften wettgemacht. So können die meisten Moosgametophyten vollständig austrocknen ohne abzusterben und Wochen bis Monate im Zustand latenten Lebens überdauern. Diesen Vorgang kann man gut beobachten, wenn man ein grünes Moospflänzchen ins Zimmer nimmt. Nach wenigen Minuten trocknen die Blätter ein und die Pflanze macht einen verwelkten, toten Eindruck. Legt man sie dann in ein Glas mit Wasser, so gewinnt sie ganz schnell wieder ihre ursprüngliche Gestalt.
-
Heterophasischer Generationswechsel (typisch für Pflanzen und viele Algen)
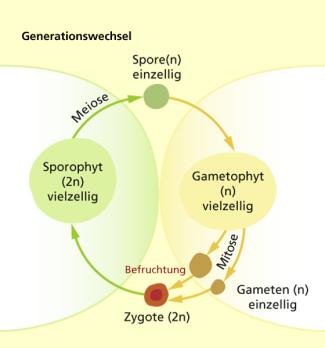

Die ersten Landpflanzen haben sich vermutlich vor ca. 450 Mill. Jahren aus schildalgenähnlichen, flächigen Grünalgen entwickelt, die am Übergang von Land und Wasser lebten und teilweise Austrocknung vertrugen. Aus der diploiden Zygote hat sich der Sporophyt entwickelt, der zunächst so ähnlich ausgesehen haben könnte wie heute noch bei Lebermoosen und auch bei Hornmoosen. Dieser Sporophyt, der ernährungsphysiologisch von Gametophyt en abhängig blieb, und bei den Moosen noch heute vom Gametophyten abhängig ist, war deutlich resistenter gegen Austrocknung und konnte auch das Austrocknen des Gametophyten längere Zeit überstehen. So konnte er auch unter trockenen Bedingungen durchhalten, bis die Sporen reif und verbreitungsfähig geworden waren. Dabei halfen ihm die Kutikula und auch die Transpiration ermöglichenden Spaltöffnungen. Eine gewisse Eigenversorgung des Sporophyten bewirkte ein entsprechendes Assimilationsgewebe, wie man es heute gut entwickelt bei den Sporophyten der Hornmoose findet. Die Höheren Landpflanzen (Gefäßpflanzen) haben sich vermutlich aus hornmoosähnlichen Vorfahren entwickelt, indem der Sporophyt sich weiter ausgestaltete und der Gametophyt schließlich als unabhängige Pflanze ganz verschwand.
Demgegenüber haben die Gametophyten der Laubmoose eine weitgehende Differenzierung und Anpassung an das Landleben erfahren. Aber auch die Gametophyten der Lebermoose sind sehr unterschiedlich gestaltet und teilweise gut an Landbedingungen angepasst.
Allen Moosen gemeinsam ist, dass sie noch nicht deutlich die drei Grundorgane der Sprosspflanzen – Sprossachse, Blätter und Wurzeln – ausgebildet haben. Im Gegensatz zu Farnpflanzen und Samenpflanzen haben ihre Vegetationskörper keine Wurzeln. Ihre Blättchen sind den Blättern der übrigen Pflanzen nicht homolog, oft bestehen sie nur aus einer Zellschicht. Der Befestigung an der Unterlage dienen einzellige (Lebermoose) oder aus einer Zellreihe (Laubmoose) bestehende Haare, die in Anlehnung an die Wurzelhaare der Sprosspflanzen als „Rhizoide“ bezeichnet werden.
Der Generationswechsel
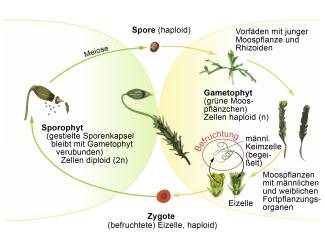
Aus den durch Meiose entstandenen Sporen eines Mooses keimt zunächst ein fädiges Protonema (Vorkeim). Es verzweigt sich und entwickelt sich zu einem grünen Filz, der sehr ähnlich aussieht, wie der Thallus (das Lager) mancher Luftalgen, z. B. der Alge Vaucheria. Hat das Protonema eine bestimmte Größe erreicht, bilden sich „Knospen“ mit zwei- oder dreischneidigen Scheitelzellen, aus denen sich die grünen Moospflänzchen entwickeln. Pro gekeimter Spore können nebeneinander sehr viele solcher Moospflänzchen entstehen. So kommt es zum typischen Rasen- oder Polsterwuchs.
Die Bedeutung der Protonemabildung liegt einmal darin, dass aus einer Spore ein ganzer Moosrasen hervorgehen kann, zum Anderen erlaubt dieses Vorstadium das Fortbestehen unter Bedingungen, die für eine Moospflanze nicht ausreichend wären, z. B. bei äußerst schlechtem Lichtangebot. Auf den Moospflänzchen entwickeln sich entweder zusammen oder auf getrennten Pflänzchen männliche und weibliche Keimzellenbehälter (Gametangien). Im ersten Fall spricht man von Einhäusigkeit (Monözie), im zweiten Fall von Zweihäusigkeit (Diözie). Die Befruchtung erfolgt durch Spermatozoiden, die über Wassertropfen zu der Eizelle in das weibliche Gametangium (Archegonium) gelangen. In jedem Archegonium ist nur eine Eizelle enthalten. Aus der befruchteten Eizelle (Zygote) entwickelt sich der Sporophyt, der zunächst als Embryo im Archegonium eingeschlossen bleibt. Während der weiteren Sporophytenentwicklung wächst zunächst das Archegonium mit, später reißt es auf, aber auf der Sporophytenkapsel bleibt der obere Teil des Archegoniums als Haube zurück.
Während sich bei den Laubmoosen zunächst der Kapselstiel streckt und dann erst die Kapsel entwickelt und ausdifferenziert, entwickeln sich bei den Lebermoosen zunächst die Kapsel, erst später der Stiel. Besonders ausgebildete „Sporophytenträger“ entwickeln sich beim Brunnenlebermoos und verwandten Arten. Die Sporophyten der Laubmoose können in jungem Zustand – wie die Hornmoossporophyten – auch chlorophyllhaltiges Assimilationsgewebe besitzen. In der Epidermis ihres Sporophyten entwickeln sie Spaltöffnungen, wie sie für alle übrigen Landpflanzen charakteristisch sind.
-
Generationswechsel bei einem Laubmoos
© Duden Learnattack GmbH

Lebensformen der Moose
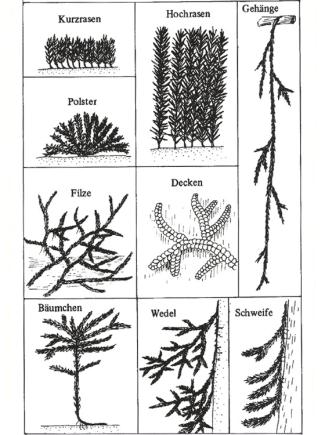
Unter „Lebensform“ einer Pflanze versteht man deren Gestalt in ihrer Beziehung zu besonderen Lebensbedingungen. Moose entstehen aus einem vielknospigen, fädigen Vorkeim (Protonema). Daraus ergibt sich, dass jeweils Verbände aus vielen Einzelpflanzen gebildet werden. Die Gestalt eines solchen Verbandes und die Wuchsform des einzelnen Moospflänzchens – Art der Verzweigung, Beblätterung, Wuchsrichtung – machen zusammen die Lebensform aus. Die vorherrschende Lebensform zeigt deutliche Beziehungen zu den ökologischen Bedingungen eines Standorts.
Hochrasen sind für feuchte bis nasse Standorte charakteristisch (Haarmützenmoos-Gattung Polytrichum; Gabelzahnmoos-Gattung Dicranum).
Kurzrasen sind charakteristisch für offene Mineralböden (Hornzahnmoos Ceratodon purpureum; Borsten-Haarmützenmoos Polytrichum piliferum).
Polster treten fast ausschließlich an freistehenden Felsen und an Baumrinde auf, aber auch an entsprechenden, von Menschen geschaffenen Standorten wie Mauern und Dächern (Mauerdrehzahnmoos Tortula muralis; Kissenmoos Grimmia pulvinata; Gattungen Orthotrichum, Zygodon, Ulota).
Als Decken bezeichnet man die Lebensform, bei der Haupt- und Nebensprosse dem Substrat aufliegen und meist mit Haftrhizoiden angeheftet sind. Sie sind typisch für glatte Borken oder Steilflächen, finden sich aber auch auf dem Erdboden. In den Tropen kommen sie als sog. epiphylle Moose auf immergrünen Blättern vor (Plattmoos Plagiothecium, Weiches Kammmoos Ctenidium molluscum, auch eine ganze Reihe von Lebermoosen: Lophocolea, Radula, Frullania).
Moosfilze bilden die Hauptvegetation moosreicher Bergwälder und Nadelforste. Hierzu zählen vor allem niederliegende Laubmoose, deren Haupt- und Seitensprosse locker durcheinander gewoben sind und einen leicht abhebbaren Belag bilden (Zypressen-Schlafmoos, verschiedene Brachythecium- und Eurhynchium-Arten, Rotstengelmoos, Grünstengelmoos).
Wedelmoose lieben schattige feuchte Standorte am Waldboden, an Baumwurzeln, in Blockfeldern und an morschen Stümpfen. Sie besiedeln im Gegensatz zu Filzen vor allem stark geneigte Unterlagen (Etagenmoos Hylocomium, Zypressenmoos Hypnum, Neckersmoos Neckera).
Als Bäumchen bezeichnet man aufrechte Moossprosse, die sich am Gipfel in einen Schopf von Seitensprossen oder von großen Blättern verzweigen (Bäumchenmoos, Fuchschwanzmoos, Rosenmoos).
Schweife bevorzugen wie Wedelmoose senkrechte Substrate. Sie können jedoch auch an trockenen Standorten, z. B. an Felsen oder auf Baumrinde, vorkommen (Eichhörnchenschwanzmoos Leucodon sciuroides).
Hängemoose kommen in unseren Breiten nicht vor. Sie bilden in tropischen und subtropischen Nebelwäldern lange Gehänge an Zweigen und Ästen der Bäume, ähnlich unseren Bartflechten. Sie können sich nur bei sehr hoher Luftfeuchtigkeit und lang anhaltender Vegetationsperiode entwickeln.
Einjährige Moose beenden ihr Wachstum mit der Ausbildung von Gametangien, und nach Reife der Sporenkapseln sterben sie ab, ohne vorher Erneuerungsknospen gebildet zu haben. Bei dieser Gruppe handelt es sich durchweg um Pioniere auf offenen Mineralböden z. B. das auf Brandstellen und mit Herbiziden kahl gespritzten Flächen häufige Wetter-Drehzahnmoos Funaria hygrometrica.
Zwischen diesen Lebensformen gibt es natürlich Übergänge. Unter Umständen kommen sogar bei ein und derselben Art verschiedene Typen vor. Das Gewelltblätterige Sternmoos Mnium undulatum z. B. besitzt sterile, wedelartige Triebe, während die fertilen Sprosse dem Bäumchentyp zuzuordnen sind.
Moose sind von den Tropen bis zur Arktis und Antarktis verbreitet. Den größten Artenreichtum und die größte Zahl unterschiedlicher Lebensformen erreichen sie in den tropischen Feuchtwäldern, Dominanz dagegen – zusammen mit Strauchflechten – in der arktischen Tundra, in Hochmooren und auf bestimmten Pionierstandorten wie Rohböden und Felsen.
-
Lebensformen der Moose

Verdunstungsschutz und Wasserspeicherung bei Moosen
Im Gegensatz zu den meisten Farn- und Samenpflanzen sind die Moospflanzen überwiegend wechselfeucht (poikilohyd). Das heißt, ihre Zellen können vorübergehende Austrocknung im Zustande latenten Lebens überdauern. Da die Gametophyten keine oder nur eine ganz schwach entwickelte Kutikula besitzen, können die grünen Moospflänzchen mit der ganzen Oberfläche Wasser aufnehmen. Bei negativem Wasserpotenzial der umgebenden Atmosphäre ist andererseits der Wasserverlust durch Verdunstung ebenfalls sehr hoch. Doch obwohl Moose grundsätzlich schlechter gegen Austrocknung geschützt sind als Farnpflanzen und Samenpflanzen haben sie doch Einrichtungen entwickelt, welche die Wasserspeicherung ermöglichen und die Wasserabgabe verzögern, um dadurch eine längere positive Fotosynthese zu gewährleisten. Dies gilt natürlich v. a. für Moose besonders trockener Standorte wie Felswände, Baumstämme, Mauern, Dächer u. ä. Das sind die typischen Standorte für Polstermoose, deren dichte Triebe kissenartige Strukturen bilden. In den dichten Polstern halten sich relativ lange Wasserreste, vor zu starker Sonneneinstrahlung und damit verbundener Erwärmung schützen oft Blättchen mit langen Glashaaren aus abgestorbenen Zellen.
Einige Moose können ihre Blättchen durch hygroskopisch arbeitende Blattgelenke bei Trockenheit an die Stämmchen anlegen. Dies könnte neben Verdunstungsschutz auch einen gewissen Strahlungsschutz bewirken.
Wieder andere Moose haben besondere Strukturen zur Wasserspeicherung entwickelt: Das Baumrinden bewohnende Lebermoos Frullania dilatata bildet auf der Unterseite seiner dicht an die Rinde anliegenden Triebe tassenartige Blättchen, in denen das Wasser besonders lange gehalten werden kann. Das Weißmoos oder Ordenskissenmoos Leucobryum ist typisch für saure, relativ trockene Waldböden. In feuchtem Zustand sieht es frisch grün, in trockenem Zustand weißlich-hellgrün aus. Dies hängt damit zusammen, dass seine Blättchen zum größten Teil aus großlumigen, abgestorbenen Zellen bestehen, die sich über Wandporen mit Wasser füllen können. Wenn dieses Wasser verdunstet, wird das Licht an den lufterfüllten Zellen reflektiert und die Polster erscheinen weißlich. Wegen dieser guten Wasserspeicherung wird das Moos gerne zur Ausgestaltung von Feuchtterrarien und für Pflanzgefäße von tropischen Orchideen verwendet. Eine besonders effektive Form der Wasserspeicherung kennt man von den Torfmoosen, durch die dadurch landschaftsprägende Geländestrukturen, die Hochmoore, entstehen konnten.
Torfmoose und Hochmoore
Moore entstehen auf wasserdurchtränkten Böden, in denen wegen Sauerstoffmangels die anfallenden Pflanzenreste nur sehr langsam abgebaut werden. Da die Produktion organischer Substanz schneller erfolgt als deren Remineralisierung, kommt es zur Ansammlung mehr oder weniger mächtiger, mineralarmer Humussubstanz (Torf). Geologisch werden Moore definiert als Böden mit einer mindestens 30 cm hohen Torfauflage, deren Gehalt an brennbarer organischer Substanz 75 % übersteigt.
Als Niedermoore oder Flachmoore bezeichnete man solche Torfmoore, die auf Wasser vom Untergrund (verlandender See, Quellwasser) angewiesen sind. Demgegenüber sind Hochmoore vom Grundwasser unabhängig und allein auf den atmosphärischen Niederschlag angewiesen. Entsprechend sind sie eng an ein bestimmtes Klima gebunden: hoher Niederschlagsüberschuss, keine zu langen Trockenperioden, mittlere Temperaturen und eine nicht zu kurze Vegetationszeit.
Voraussetzung für die Hochmoorbildung ist die Ansiedelung der Torfmoose (Gattung Sphagnum), eine Unterklasse der Laubmoose mit ganz besonderen morphologischen, physiologischen und ökologischen Eigenschaften. Die Sphagnum-Polster saugen sich wie Schwämme voll mit Wasser. Bei feuchtem Klima können solche Polster und Rasen immer höher über den Grundwasserspiegel emporwachsen. Sie zersetzen sich nur langsam und die unteren Schichten der Polster gehen allmählich in Sphagnum-Torf (Weißtorf) über, wobei als Zersetzer vor allem Pilze eine wichtige Rolle spielen. In der Rhön konnte ein Torfzuwachs von 10 cm pro Jahrhundert festgestellt werden, in feuchteren und milderen Klimaten Westeuropas mag dieser Wert sogar noch deutlich höher liegen. Im Laufe der Jahrtausende sind so mächtige, Regenwasser getränkte Torfschilde entstanden, die sich uhrglasförmig mehrere Meter über das Relief erheben. Für die wachsenden Torfmoose an der Oberfläche ist es wichtig, dass sie weder zu lange vom Wasser überflutet sind noch zu lange trocken liegen. Der Wasserstand sollte 10 bis 15 cm unterhalb der Sphagnum-Köpfchen liegen. Aus den Rändern des Torfschildes sickert saures, nährstoffarmes Wasser und sammelt sich in einem Randsumpf, dem sogenannten Lagg. Das aus dem Moor ablaufende Wasser schafft sich mit der Zeit radiär verlaufende Abflussrinnen, sogenannte Rüllen. Auch kleinere Seen, sogenannte Kolke oder Blänken, können sich auf der Moorfläche bilden.
Hohe Wasserspeicherfähgikeit
Sphagnen können das bis zu 20fache ihres Eigengewichts an Wasser festhalten. Diese Fähigkeit verdanken sie vor allem besonderen Wasserspeicherzellen in den Blättchen und in der Rinde des Stämmchens. Es sind tote, großlumige Zellen, deren Außenwände mit Poren versehen sind. Durch diese Poren können sie durch Kapillarkräfte sehr rasch Wasser einsaugen. Bei Verdunstung schützen besondere Spiralversteifungen die Wasserspeicherzellen vor dem Kollabieren. Außerdem wirken sich die dichte Stellung der quirlig angeordneten Seitenzweige sowie der Polsterwuchs günstig auf das Wasserhaltevermögen aus.
Kapillare Wasserleitung
Neben der Speicherung können die verschiedenen Kapillarräume des Sphagnum-Polsters auch der aufwärts gerichteten Wasserleitung dienen. Besonders wirksam ist hierbei die dochtartige Wirkung von Seitenzweigen, die eng anliegend am Hauptstamm herab laufen.
Kationenaustausch
Ein besonderer Ionen-Austausch-Mechanismus gestattet den Sphagnum-Pflänzchen, die wenigen Kationen, die im Hochmoorwasser gelöst sind, im Austausch gegen H+-Ionen ,,herauszufangen". Die Torfmoospolster stellen dadurch Kationen-Austauschsäulen dar, die sich im Wechsel von Regen und aufsteigendem Wasserstrom bei Verdunstung selbst regenerieren. Eine wichtige Rolle bei diesem Austausch spielen Bestandteile der Zellwände, doch sind die lebenden Zellen in den Kationen-Kreislauf eingebunden und können dadurch mit K+- und NH4 +-Ionen versorgt werden. Nebeneffekt des Ionenfangs ist die Ansäuerung des umgebenden Wassers. Diese Ansäuerung nimmt mit dem Wachstum der Sphagnen zu. Je mehr Sphagnen in einem Wasservolumen stehen, desto saurer ist dieses Wasser. Hierdurch werden fast alle anderen Pflanzen ausgeschaltet.
Hohe Wasserspeicherfähgikeit
Sphagnen können das bis zu 20fache ihres Eigengewichts an Wasser festhalten. Diese Fähigkeit verdanken sie vor allem besonderen Wasserspeicherzellen in den Blättchen und in der Rinde des Stämmchens. Es sind tote, großlumige Zellen, deren Außenwände mit Poren versehen sind. Durch diese Poren können sie durch Kapillarkräfte sehr rasch Wasser einsaugen. Bei Verdunstung schützen besondere Spiralversteifungen die Wasserspeicherzellen vor dem Kollabieren. Außerdem wirken sich die dichte Stellung der quirlig angeordneten Seitenzweige sowie der Polsterwuchs günstig auf das Wasserhaltevermögen aus.
Kapillare Wasserleitung
Neben der Speicherung können die verschiedenen Kapillarräume des Sphagnum-Polsters auch der aufwärts gerichteten Wasserleitung dienen. Besonders wirksam ist hierbei die dochtartige Wirkung von Seitenzweigen, die eng anliegend am Hauptstamm herab laufen.
Kationenaustausch
Ein besonderer Ionen-Austausch-Mechanismus gestattet den Sphagnum-Pflänzchen, die wenigen Kationen, die im Hochmoorwasser gelöst sind, im Austausch gegen H+-Ionen ,,herauszufangen". Die Torfmoospolster stellen dadurch Kationen-Austauschsäulen dar, die sich im Wechsel von Regen und aufsteigendem Wasserstrom bei Verdunstung selbst regenerieren. Eine wichtige Rolle bei diesem Austausch spielen Bestandteile der Zellwände, doch sind die lebenden Zellen in den Kationen-Kreislauf eingebunden und können dadurch mit K+- und NH4 +-Ionen versorgt werden. Nebeneffekt des Ionenfangs ist die Ansäuerung des umgebenden Wassers. Diese Ansäuerung nimmt mit dem Wachstum der Sphagnen zu. Je mehr Sphagnen in einem Wasservolumen stehen, desto saurer ist dieses Wasser. Hierdurch werden fast alle anderen Pflanzen ausgeschaltet.

