Erregbarkeit tierischer Zellen
Bei Tieren haben sich für Erregung und Erregungsleitung spezielle Zell- und Gewebetypen entwickelt:
Sinneszellen (Rezeptorzellen) sind spezialisierte Nervenzellen bzw. Nervenzellanteile für die Reizaufnahme.
Primäre Sinneszellen sind Reiz aufnehmende Zellen, deren Zellkörper unmittelbar in den erregungsleitenden Neuriten übergeht. Sie kommen vor allem in den Sinnesorganen wirbelloser Tiere vor.
Sekundäre Sinneszellen sind modifizierte Epithelzellen, die von Endigungen sensibler Neurite umschlossen werden. Dieser Sinneszelltyp wurde bisher nur bei Wirbeltieren nachgewiesen.
Freie Nervenendigungen werden durch feinste Verzweigungen der sensiblen Neurite gebildet. Sie sind vor allem bei Wirbeltieren als Rezeptoren des Tast- und Temperatursinnes verbreitet.
Nervenzellen (Neuronen) sind für die Prozesse der Informationsübertragung und Informationsverarbeitung spezialisierte Zellen. Während Stoffwechselvorgänge im Zellkörper (Soma) speziell in den Mitochondrien ablaufen, dienen die Fortsätze der Erregungsleitung. Über Dendrite werden Erregungen zugeführt, Neurite (Axone) sichern die Erregungsweiterleitung.
Neurite, die die Erregung von den Sinnesorganen zum Rückenmark und Gehirn leiten, sind sensible oder afferente Neurite. Solche Neurite, die die Erregung vom Gehirn und Rückenmark an die Muskeln leiten, sind motorische oder efferente Neurite. Die seitlichen Verzweigungen des Neuriten werden als Kollaterale bezeichnet.
Verbindungen zwischen den Nervenzellen untereinander und zu anderen Zellen (z. B. Muskelzellen) werden über Synapsen, bläschenförmige Erweiterungen an den Enden der Neuriten und Dendriten, hergestellt. In den Synapsen findet die Erregungsübertragung zwischen zwei Zellen statt.
Viele Neurite sind von einer Markscheide (lipid- und eiweißreiche Myelinhülle), umgeben, die aus den schwannschen Zellen gebildet wird.
Die Markscheide wirkt wie die Isolation eines Kabels und wird in Abständen von etwa 1 bis 2 mm durch die ranvierschen Schnürringe unterbrochen. Sie spielen bei der Erregungsleitung eine wesentliche Rolle.
LOUIS ANTE RANVIER (1835–1922) war als Histologe in Paris an der Entdeckung der kontinuierlich auftretenden Einschnürungen der Markscheide beteiligt, daher die Bezeichnung ranvierscher Schnürring. THEODOR SCHWANN (1810–1882) untersuchte tierische Zellen. Er entdeckte u.a. Zellen, die die Markscheide ausbilden (schwannsche Zellen).
In den Muskelzellen werden die Erregungen der Neuronen in Bewegung umgesetzt. Man unterscheidet glatte Muskelzellen, quer gestreifte Muskelzellen und die Herzmuskelzellen.
Glatte Muskelzellen sind lang gestreckte spindelförmige Zellen mit einer durchschnittlichen Länge von 0,05 bis 0,2 mm. Sie haben einen Zellkern und wenig strukturiertes Cytoplasma, das bei Muskelzellen als Sarkoplasma bezeichnet wird. Im Sarkoplasma befinden sich kontraktionsfähige parallel verlaufende fädige Strukturen, die Myofibrillen. Glatte Muskelzellen bilden die Muskulatur der inneren Organe von Wirbeltieren und die „Haltemuskulatur“ bei einigen Wirbellosen (z. B. Schalenschließmuskel der Muschel). Sie entwickeln einen guten Muskeltonus (Daueranspannung) ohne größeren Energieverbrauch.
Quer gestreifte Muskelzellen bilden bei Wirbeltieren und Arthropoden die Skelettmuskulatur und kommen auch im Schirmrand von Quallen und im Schlundkopf der Gliederwürmer vor. Sie enthalten mehrere Zellkerne. Im Zellplasma (Sarkoplasma) befinden sich als kontraktile Elemente einige 100 Myofibrillen. Die kleinste Einheit einer Myofibrille ist ein ca. 2 mm langer Sarkomer, der beidseitig von der Z-Membran begrenzt ist. Im Lichtmikroskop kann man helle und dunkle Bänder unterscheiden, die durch die parallele Lage der Myofibrillen zustande kommen.
Die gesamte Muskelfaser ist von einem weitverzweigten Röhrensystem, dem sarkoplasmatischen Retikulum und dem T-System (transversales System, Einstülpungen der Zellmembran) durchzogen. Es ist für die Erregungsübertragung und die -Speicherung und -Freisetzung wichtig. Quer gestreifte Muskeln kontrahieren schnell, sie werden vom ZNS über die motorischen Endplatten innerviert.
Herzmuskelzellen ähneln in ihrem Aufbau den quer gestreiften Muskelzellen. Sie bilden ein Netzwerk, und nur die Glanzstreifen deuten den zellulären Aufbau an. Die ständig arbeitenden Herzmuskelzellen haben einen hohen Sarkoplasmaanteil. Herzmuskelzellen werden über Erregungsbildungszentren des Herzens in ihrer Funktion koordiniert und durch das vegetative Nervensystem beeinflusst.
Werden diese speziellen, erregbaren Zellen gereizt, so verändert sich ihr Ruhepotenzial. In den Nerven- und Muskelzellen entsteht das Aktionspotenzial und in den Rezeptorzellen das Rezeptorpotenzial.
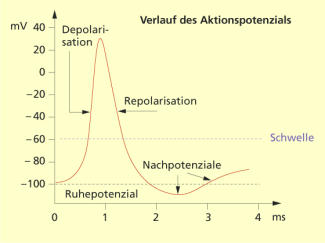
Das Aktionspotenzial (AP) ist durch eine sehr schnelle Depolarisation der Membran gekennzeichnet, bei der ein Ladungsüberschuss von +30 mV auf der Membraninnenseite gemessen werden kann. Im Anschluss kommt es zu einer raschen Repolarisation.
Diese Potenzialänderungen werden durch die spannungsabhängigen Ionenkanäle (elektrogene Pumpen) möglich. Bei Depolarisation der Membran über einen bestimmten Wert (Schwellenwert) öffnen sich die -Kanäle. können die Membran passieren, wodurch die positive Ladung der Membranaußenseite geringer wird. Das führt zur Öffnung weiterer -Kanäle und zur explosionsartigen Veränderung der Spannungsverhältnisse an der Membran. Die Außenseite ist jetzt negativer als die Innenseite geladen. Nun schließen sich die -Kanäle und die -Kanäle öffnen sich. werden aus der Zelle transportiert, bis das Ruhepotenzial wieder erreicht ist (Repolarisation). Während der Repolarisationsphase sind die Nerven- und Muskelzellen nicht wieder erregbar, sie befinden sich in der Refraktärphase .
-
Verlauf des Aktionspotenzials
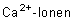

Die Zeitdauer des Aktionspotenzials variiert in den erregbaren Zellen. Sie ist an Axonen am kürzesten (1 bis 2 ms) und an Herzmuskelzellen am längsten (bis 200 ms).
Aktionspotenziale (AP) folgen der Alles- oder Nichtsregel, d. h., ist das Schwellenpotenzial überschritten, haben sie – unabhängig von der Reizstärke – dieselbe Größe. Die Information über die Reizstärke wird durch die Frequenz der aufeinanderfolgenden AP codiert.
Während Nerven und Muskeln durch Depolarisation benachbarter Zellen und im Experiment durch elektrische Reizung erregt werden, nehmen Rezeptorzellen die Reize der Umwelt (z. B. optischer Reiz, akustischer Reiz, mechanischer Reiz, chemischer Reiz) auf, reagieren mit Potenzialänderungen (Rezeptorpotenzial) und bewirken das Aktionspotenzial in den anschließenden erregungsleitenden Nervenfasern.
Das Rezeptorpotenzial ist von der Reizstärke abhängig, je stärker der Reiz, umso höher ist die Amplitude der De- bzw. Hyperpolarisation in den Rezeptorzellen. Eine Hyperpolarisation liegt vor, wenn eine Verstärkung des negativen Spannungsbereiches über den Wert des Ruhepotenzials hinaus geht.
Die meisten Rezeptoren reagieren mit einer Depolarisation, d. h., mit einer erhöhten -Permeabilität der gereizten Membran, die in der gesamten Zelle noch verstärkt wird. Bei adäquaten Reizen genügt schon eine geringe Reizstärke, um diese Reaktion auszulösen.
Lichtsinneszellen reagieren bei Belichtung mit Hyperpolarisation, d. h., die -Permeabilität verringert sich bei Reizung. Diese ersten Umwandlungen des Reizes in einen zelleigenen Prozess (Membranpotenzialänderungen) bezeichnet man als Transduktion.
Das Rezeptorpotenzial löst an den erregungsleitenden Nervenfasern, die mit der Rezeptorzelle in Verbindung stehen, ein Aktionspotenzial aus. Diesen Vorgang bezeichnet man als Transformation des Rezeptorpotenzials. Die Frequenz der Aktionspotenziale ist von der Amplitude des Rezeptorpotenzials abhängig.
-
Zeitverlauf der Aktionspotenziale

Suche nach passenden Schlagwörtern
- sarkoplasmatisches Retikulum
- Reize
- Muskelfaser
- Synapsen
- sensible
- Neurite
- Depolarisation
- Aktionspotenzial
- Glatte Muskelzellen
- Kollaterale
- Amplitude des Rezeptorpotenzials
- Markscheide
- Sinneszellen
- Soma
- Erregungsleitung
- Dendrite
- Sarkomer
- Skelettmuskulatur
- motorische
- Neuronen
- Myofibrillen
- Sarkoplasma
- Zellkörper
- Transformation
- Nervenzellen
- Erregbarkeit
- efferente
- Refraktärphase
- Erregung
- Herzmuskelzellen
- afferente
- Lichtsinneszellen
- schwannschen Zellen
- Myelinscheide
- Axone
- Transduktion
- ranvierschen Schnürringe
- Sinnesorgane
- Quer gestreifte Muskelzellen

